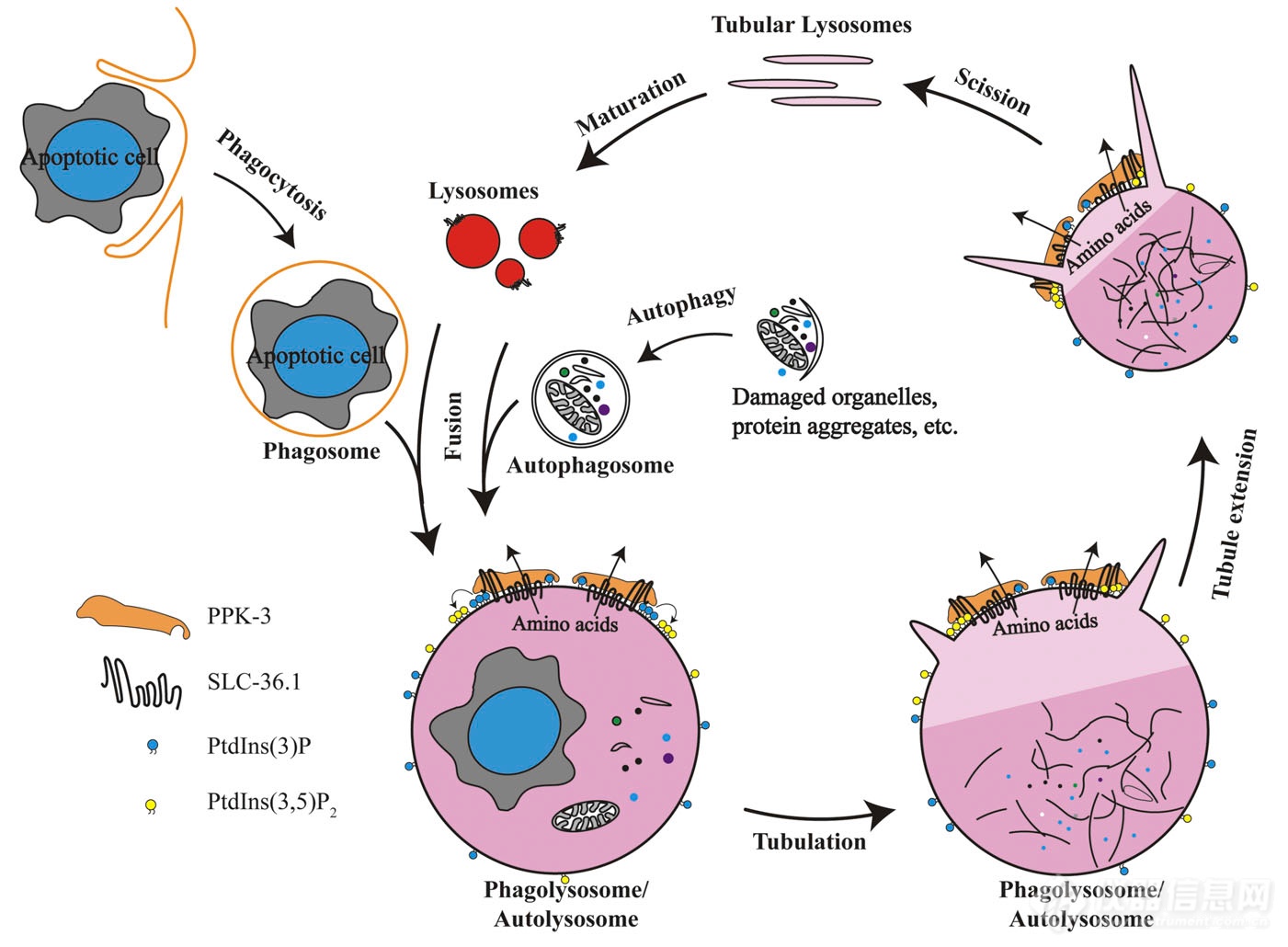
溶酶体在细胞内发挥重要功能,此次整理分享的文章研究的是溶酶体的再形成,文章作者以线虫胚胎发育过程中细胞尸体的清除为研究模型,发现细胞尸体被吞噬后溶酶体的再形成受SLC-36.1和PPK-3调控,具体内容如下:
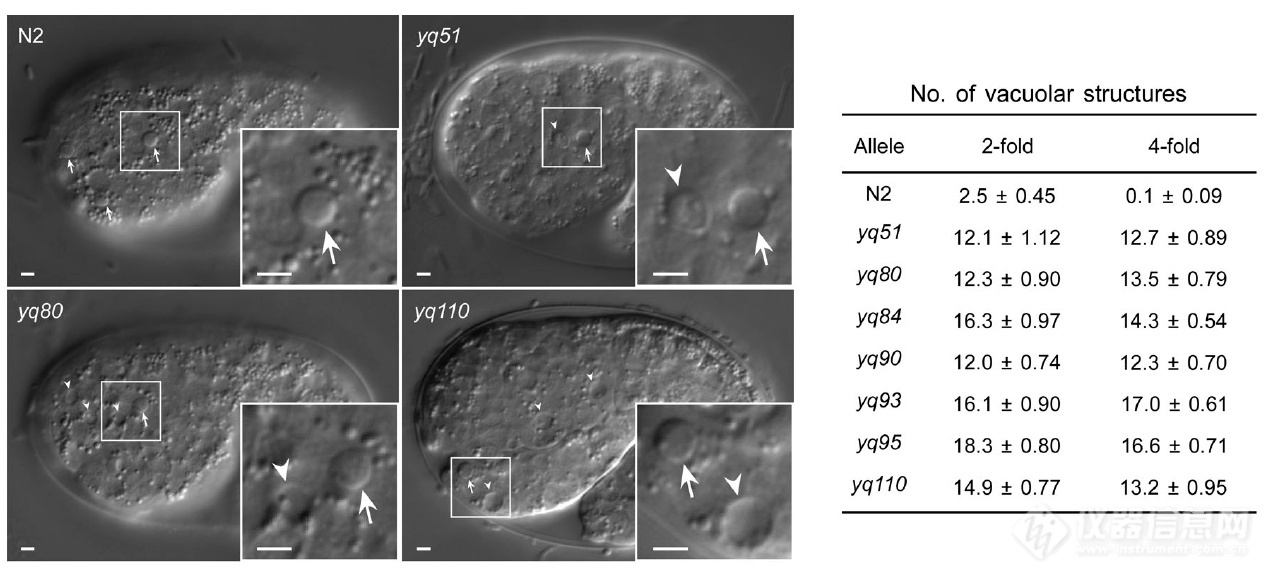
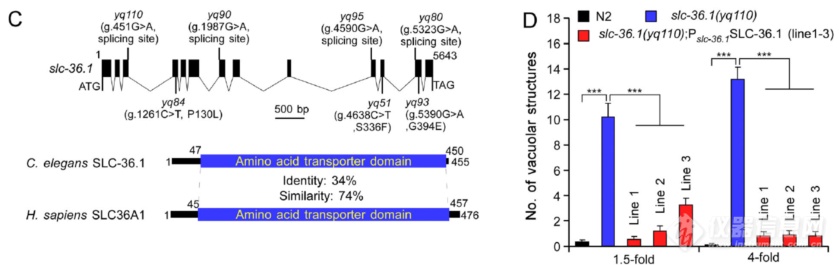
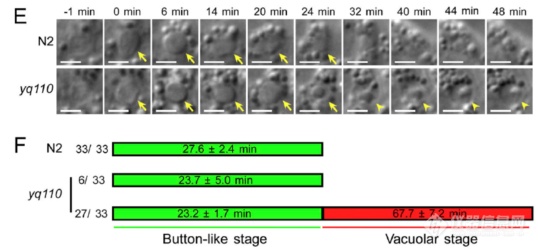
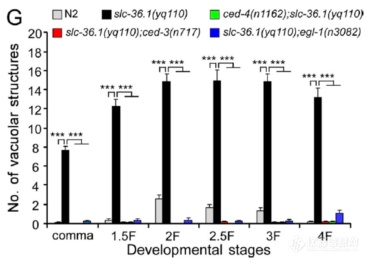
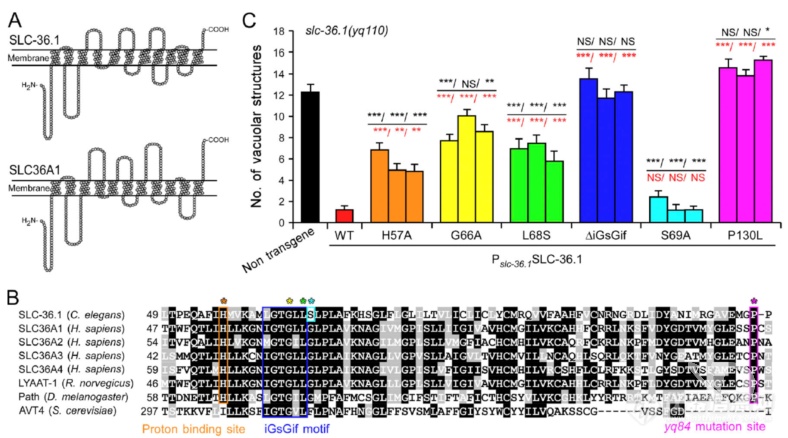
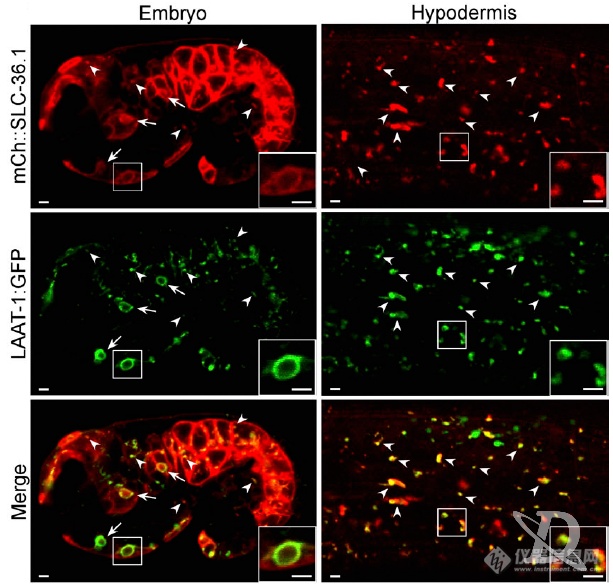
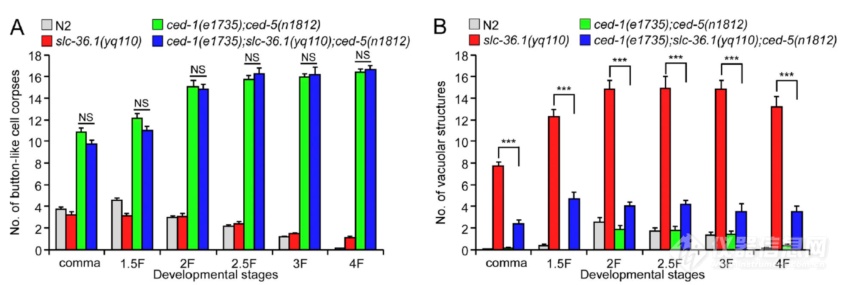
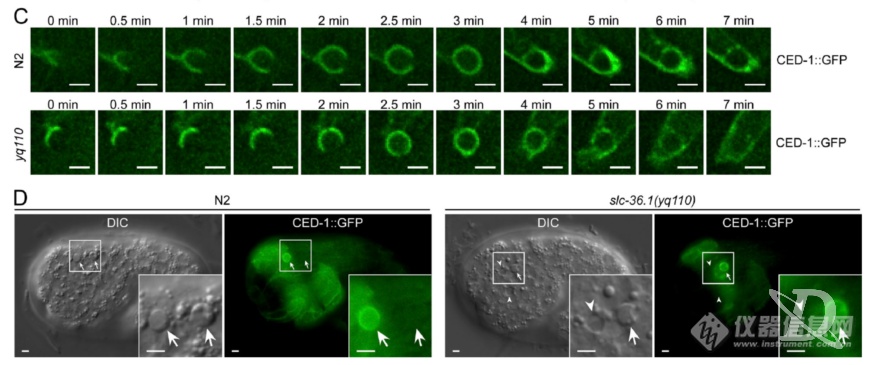
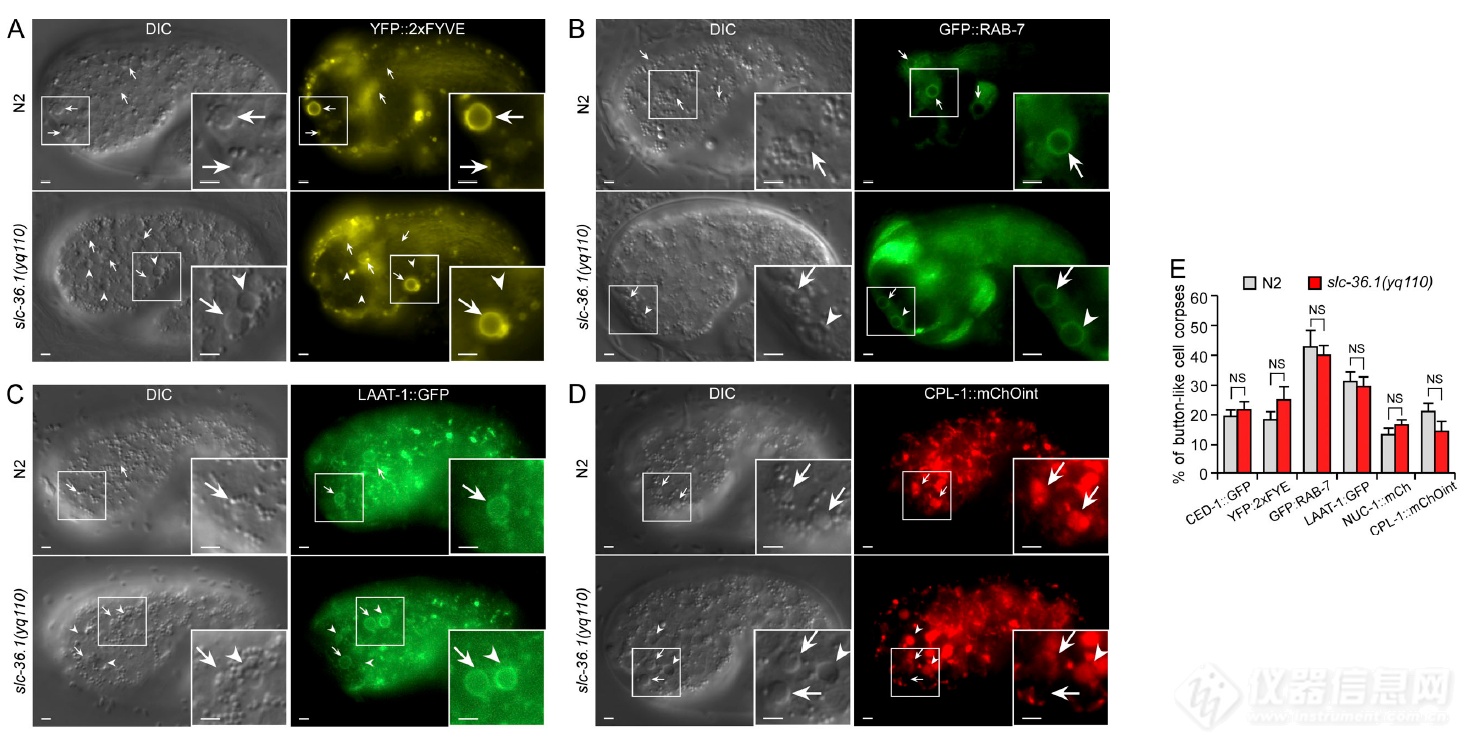
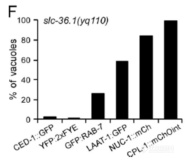
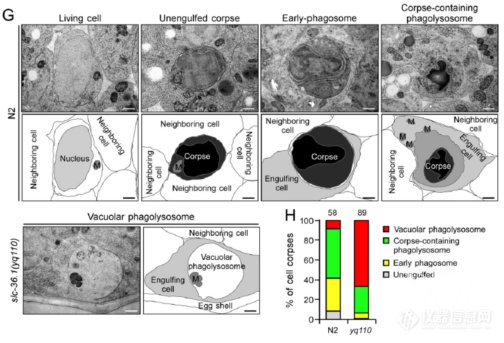
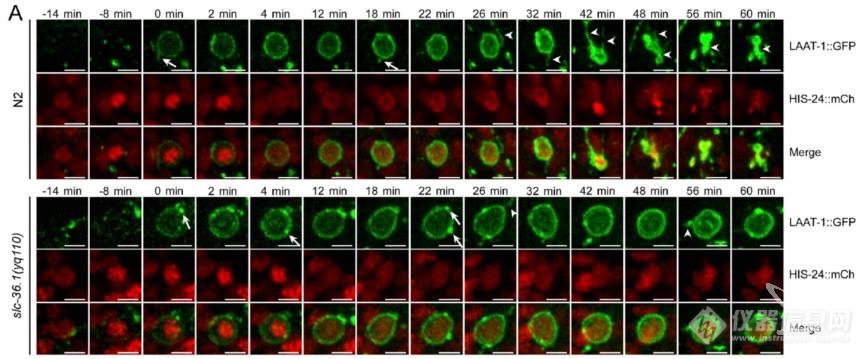
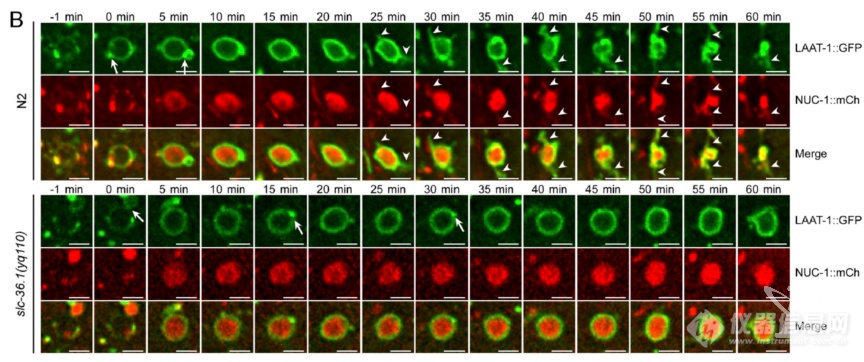
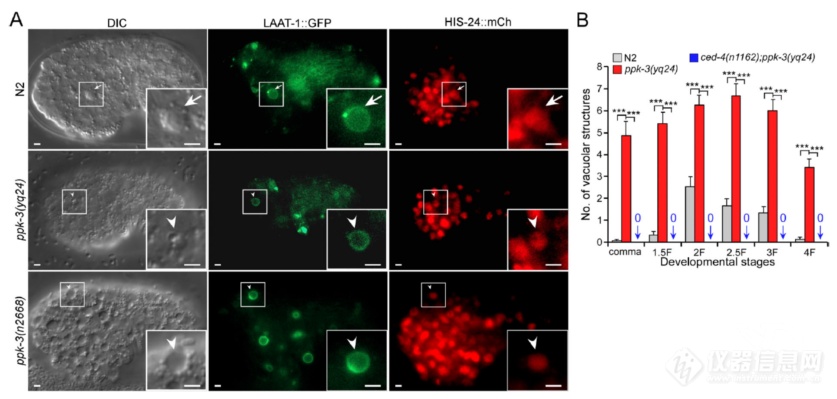
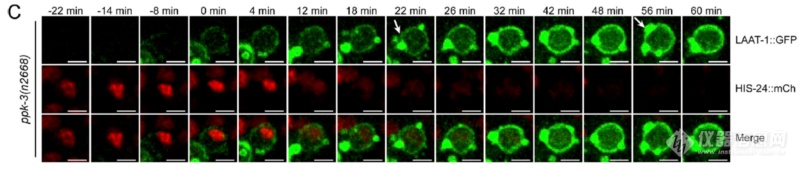
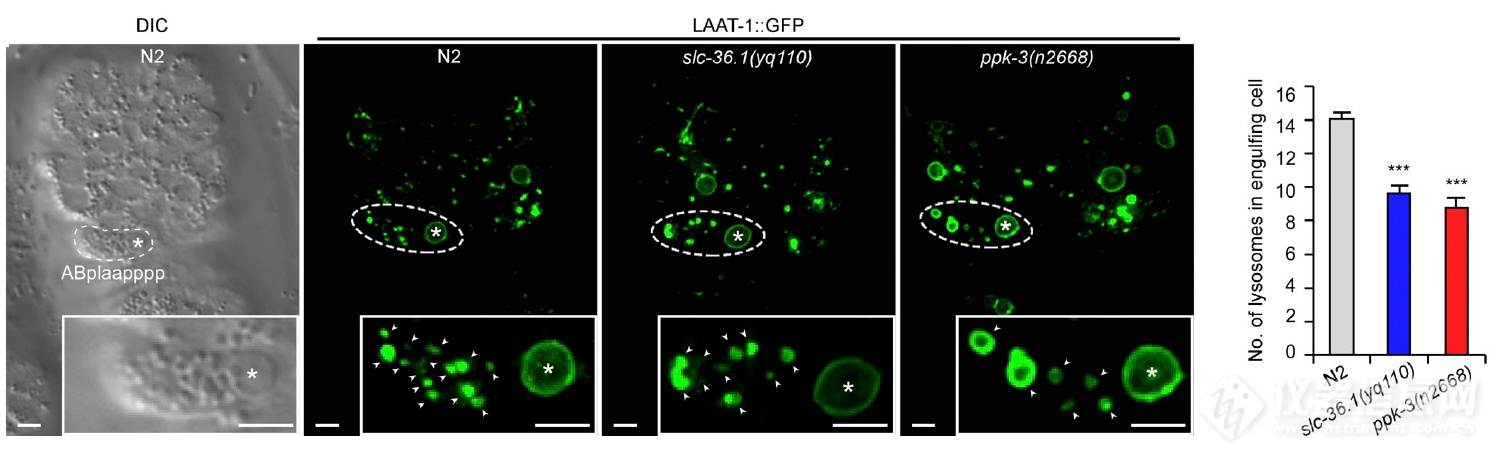
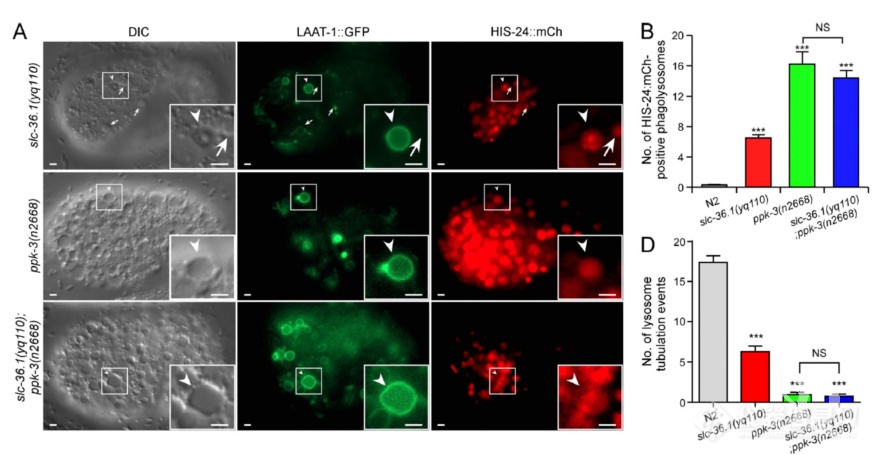
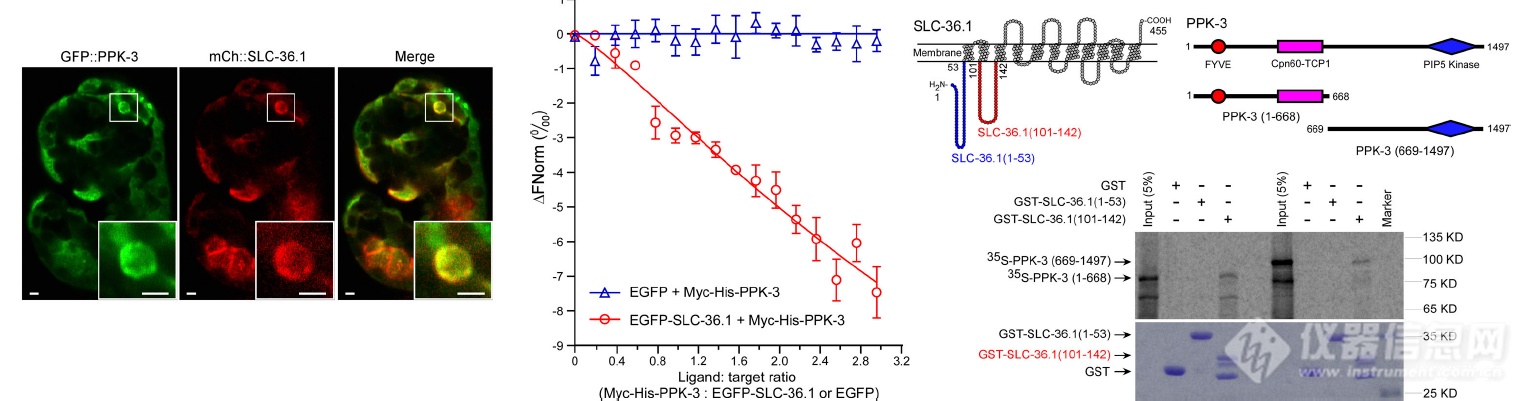
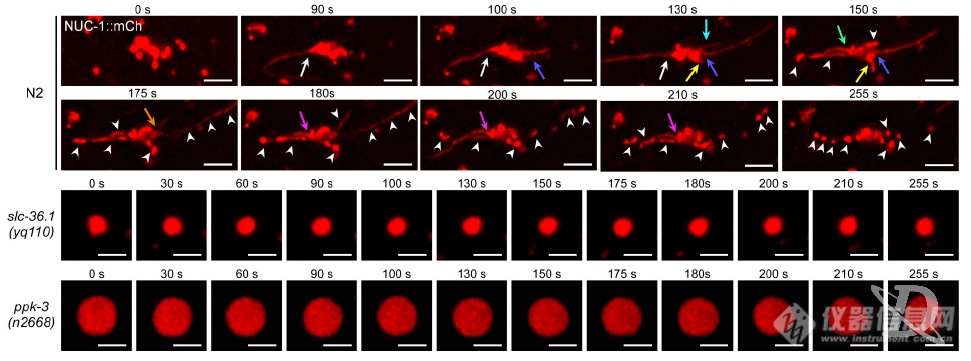
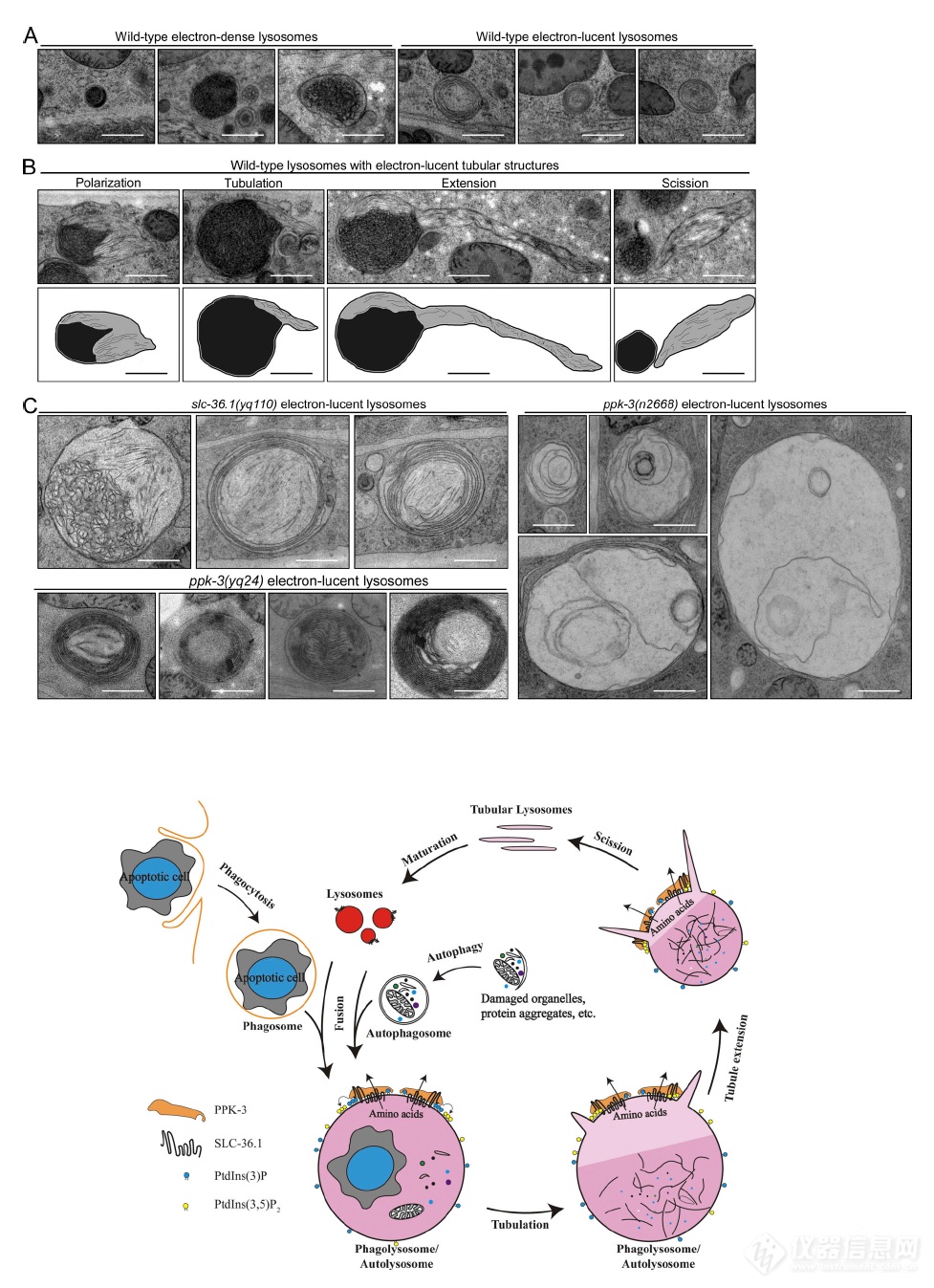
文章作者提到过自噬溶酶体形成后溶酶体再生成的调节已经研究清楚了,吞噬溶酶体形成后溶酶体再生成无人研究。之于我们,工具、模型其实就那么多;之于细胞,细胞器、功能分子就那么多,底层的逻辑其实变化不会太大,差异就在于对这些基本点的把握,工具、模型是open的,只是掌握的水平不同̷̷
Qiwen Gan, Xin Wang, Qian Zhang, Qiuyuan Yin, Youli Jian, Yubing Liu, et al. The amino acid transporter SLC-36.1 cooperates with PtdIns3P 5-kinase to control phagocytic lysosome reformation [J]. J. Cell Biol. 2019.
想了解更多CNS级期刊更新内容,请关注我们的公众号,常有更新哦,也可加笔者微信交流:qianle522568
【Webinar 预告】肝脏类器官研究进展及应用分享

【邀请函】第七届酶标仪应用及软件操作高级培训班北京站开班,邀您报名!

【订购有礼】限时狂欢!电商专享商品上线,每单必得惊喜好礼!

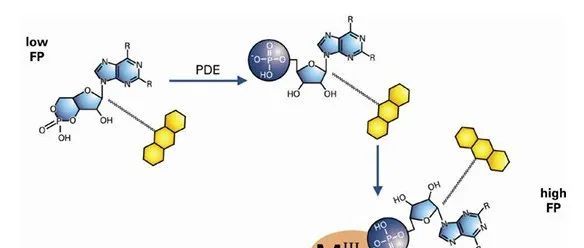
酶标仪科普专栏丨第二期:荧光偏振(FP)介绍及应用






相关产品
上海美谱达UV-3200扫描型紫外/可见分光光度计
Leica徕卡DM750M正置显微镜
希森美康ExoCounter 全自动外泌体分析仪
智能化共聚焦高内涵成像分析系统
比利时 Bioruptor plus非接触式全自动超声破碎仪
DM3000德国徕卡leica生物显微镜
徕卡生物倒置显微镜LeicaDMi1
希森美康ExoCounter 全自动外泌体分析仪
梅特勒托利多pH 复合电极 LE438
Diagenode全自动核酸剪切仪Megaruptor ®3
化学发光成像仪Amersham ImageQuant 800
BIORUPTOR超声波细胞破碎仪new pico比利时Diagenode
美国博勒飞AMETEK Brookfield CTX质构仪
美国思拓凡 cytiva go蛋白纯化系统
ATS超高压均质机AH-D400

关注
拨打电话

留言咨询





