
应激颗粒SG(stress granule)是很典型的相变产物,SG内有各种蛋白及核酸,蛋白G3BP1是SG内重要的组分,影响着相变液滴的形成,此次分享的文章研究的就是G3BP1,具体内容如下:

观察到G3BP1相变

看相变细节:FRAP,RNA参与相变
 纯化出G3BP1,细胞外看G3BP1相变行为
纯化出G3BP1,细胞外看G3BP1相变行为

调节G3BP1二聚的NTF结构域、精氨酸多的正电性结构域RG影响相变(没用RNA,用的PEG)

细胞受到刺激后产生的应激颗粒(SG)包含多种蛋白质,作者在细胞外先构建了G3BP1相变液滴,而后看其他蛋白的“参与”情况
 Caprin-1自己不相变,和GBP1一起则相变,且可通过增强G3BP1对RNA的结合能力(EMSA技术评估蛋白和RNA的结合能力)降低GBP1的相变阈值
Caprin-1自己不相变,和GBP1一起则相变,且可通过增强G3BP1对RNA的结合能力(EMSA技术评估蛋白和RNA的结合能力)降低GBP1的相变阈值

看另一个蛋白对G3BP1相变液滴的影响:两种形式的Ubc9(WT为野生型,ts为折叠错误的突变型),两种形式的Ubc9在溶液中扩散能力没有差异,折叠错误的突变型ts能够进入G3BP1的相变液滴,使其硬化(融合减弱、内部分子运动受限) - 液滴时可调节生化反应,硬化了往往会影响细胞功能,引起疾病

G3BP1自己不相变,和RNA一起才发生相变,所以作者看看什么样的RNA才能引起相变:长的、未折叠的RNA才能有效触发相变

G3BP1上不同的结构域影响着它和RNA的结合能力,通过和RNA一起相变可以抑制RNA自己不可逆的纠错在一起

IDR结构域对相变的影响还没讨论,用各种算法分析后发现IDR结构域呈负电性,G3BP1和RNA一起才能相变,RNA是负电性的,所以负电性的IDR结构域对相变应该有负调节作用,IDR的负电性主要来源于酸性氨基酸,通过调低pH,抑制-COOH裸露出负电可促进相变

调pH“关掉”负电基团后又把负电基团给敲掉了,敲掉之后相变增强

发散一下,G3BP1上的磷酸化位点会带来负电性的磷酸基团,对相变也是负调节作用

更进一步的看负电荷结构域,G3BP1分子内同时有正负电结构域,分子内结构域会不会存在相互作用呢,软件模拟发现带负电的酸性结构域在空间上的确和正电结构域RG相互接近,暗示G3BP1呈压缩构象,通过动态光散射技术(DLS)测得流体动力学半径反映构象状态,发现盐浓度、结构域、pH均会影响G3BP1的构象,相较压缩构象,舒展构象下G3BP1和RNA结合能力更强,更利于相变 - 挖出了蛋白构象对于相变的影响

继续挖掘DLS的数据,除了看到二聚形式的G3BP1,还看到了一小部分寡聚的G3BP1,寡聚形式的G3BP1会因为盐浓度的升高而减少;RNA能够通过和RG结构域互作促进G3BP1的寡聚化

基于对G3BP1研究得到的数据,作者构建了一个G3BP1、RNA互作的模型,用于预测两者混合后所能产生的结构:当G3BP1和短RNA混合后G3BP1可通过RNA寡聚,但并不足以发生相变

当G3BP1和长RNA混合后,蛋白和RNA纠缠在一起成簇,这种簇状结构发挥类似胶水的作用把RNA链交联成网状

模拟是一回事,实测是另一回事,作者实测:G3BP1和RNA一起可以形成相变液滴,通过GFP浓度-荧光标准曲线可计算出相变液滴内外G3BP1的浓度;G3BP1在相变液滴内是如何分布的呢,通过折射率计算浓度得到相变液滴的质量浓度大概在65mg/mL,而完全由G3BP1组成的液滴质量浓度可达400mg/mL,暗示G3BP1并没有完全充满液滴,而是和作者构建的模型类似,充当的是交联RNA的“胶水”,电镜下确实观察到了G3BP1离散分布

作者的构建的模型说明G3BP1聚簇后会形成多价的RG,可交联RNA,发生相变;G3BP2比G3BP1 RG结构域上有更多的正电性精氨酸,基于模型应该更易发生相变,实际做一下果然如此;DLS在G3BP2上观察到了和G3BP1相同的二聚和寡聚现象

最后来个模式图:未受到刺激时细胞内G3BP蛋白成压缩构象,受到刺激后mRNA释放出来,竞争性的抑制G3BP分子内部正负电结构域的互作,使G3BP变为舒展构象,在RNA的作用下寡聚,发挥“胶水”的作用,将RNA交联到一起,形成相变液滴
这篇文章把已然被发现的SG做细了,细到蛋白构象水平,勾勒出相变液滴内的分子行为。怎么细的?出发点是什么?是电性,在“正负相吸”的指导思想下除了挖分子间的行为,还挖出了分子内的行为,简单却又精彩……
Jordina Guille′ n-Boixet, Andrii Kopach, Alex S. Holehouse, et al. RNA-Induced Conformational Switching and Clustering of G3BP Drive Stress Granule Assembly by Condensation [J]. Cell, April 16, 2020.

主题:CNS前沿文献追踪–RNA通过改变G3BP1构象驱动相变进行
时间:4月29日14:00-15:00
主讲人:于博士
点击链接直接加入会议:
https://meeting.tencent.com/s/5lhqZC450d52
会议 ID:670 001 708
参考文献:
Jordina Guille′ n-Boixet, Andrii Kopach, Alex S. Holehouse, et al. RNA-Induced Conformational Switching and Clustering of G3BP Drive Stress Granule Assembly by Condensation [J]. Cell, April 16, 2020.
【CNS前沿文献追踪】Sup35在应激条件下通过相变保护细胞
【CNS前沿文献追踪】SIM超分辨看DNA损伤修复过程中γH2AX
【CNS前沿文献追踪】Amo1蛋白调节异染色质核内定位及基因沉默
【CNS前沿文献追踪】组蛋白甲基化reader ZCWPW1调节减数分裂
【CNS前沿文献追踪】用SIM观察减数分裂过程中DNA修复、联会、交叉互换
【CNS前沿文献追踪】提高肿瘤免疫疗法敏感度的新靶点ADAR1
【CNS前沿文献追踪】调节DC细胞抗原提呈的mRNA甲基化读取蛋白YTHDF1
【CNS前沿文献追踪】WDFY2通过VAMP3抑制基质金属蛋白酶再循环
【CNS前沿文献追踪】用电转导入荧光标记抗体进行活细胞超分辨成像
【CNS前沿文献追踪】用SIM观察减数分裂过程中DNA修复、联会、交叉互换


【Webinar 预告】肝脏类器官研究进展及应用分享

【邀请函】第七届酶标仪应用及软件操作高级培训班北京站开班,邀您报名!

【订购有礼】限时狂欢!电商专享商品上线,每单必得惊喜好礼!

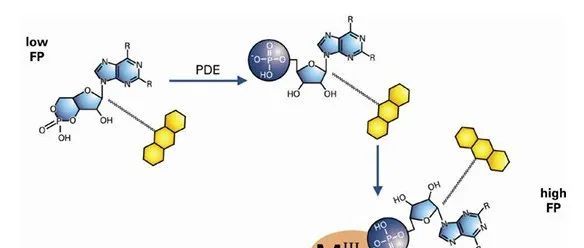
酶标仪科普专栏丨第二期:荧光偏振(FP)介绍及应用





相关产品
上海美谱达UV-3200扫描型紫外/可见分光光度计
Leica徕卡DM750M正置显微镜
希森美康ExoCounter 全自动外泌体分析仪
智能化共聚焦高内涵成像分析系统
比利时 Bioruptor plus非接触式全自动超声破碎仪
DM3000德国徕卡leica生物显微镜
徕卡生物倒置显微镜LeicaDMi1
希森美康ExoCounter 全自动外泌体分析仪
梅特勒托利多pH 复合电极 LE438
Diagenode全自动核酸剪切仪Megaruptor ®3
化学发光成像仪Amersham ImageQuant 800
BIORUPTOR超声波细胞破碎仪new pico比利时Diagenode
美国博勒飞AMETEK Brookfield CTX质构仪
美国思拓凡 cytiva go蛋白纯化系统
ATS超高压均质机AH-D400

关注
拨打电话

留言咨询





