原创与否转贴线粒体功能状态和不少疾病的密切相关,线粒体膜电位(MMP)则是反映细胞内线粒体功能状态的重要参数之一。本人整理一下线粒体膜电位测量方法,包括主要测量仪器和常用荧光探针,欢迎补充讨论。常用测量仪器:(1)普通荧光显微镜;(2)激光扫描共聚焦显微镜;(3)流式细胞仪。常用荧光探针:JC-1,DioC6,mitocapture,罗丹明123,TMRM等。JC-1(也称CBIC2(3))是一种广泛用于检测线粒体膜电位(mitochondrial membrane potential)△Ψm 的理想荧光探针。可以检测细胞、组织或纯化的线粒体膜电位。在线粒体膜电位较高时,JC-1聚集在线粒体的基质(matrix)中,形成聚合物(J-aggregates),可以产生红色荧光;在线粒体膜电位较低时,JC-1不能聚集在线粒体的基质中,此时JC-1为单体(monomer),可以产生绿色荧光。这样就可以非常方便地通过荧光颜色的转变来检测线粒体膜电位的变化。JC-1单体可采用488或514nm激光激发,发出绿色荧光波长为529nm左右;JC-1聚合物(J-aggregates)的最大激发波长为585nm,发出红色波长为590nm。罗丹明123(Rhodamine 123, Rh123)是一种可透过细胞膜的阳离子荧光染料,在正常细胞中能够依赖线粒体跨膜电位进入线粒体基质,荧光强度减弱或消失。在细胞凋亡发生时,线粒体膜完整性破坏,线粒体膜通透性转运孔开放,引起线粒体跨膜电位(ΔΨm) 的崩溃,Rh123 重新释放出线粒体,从而发出强黄绿色荧光,通过荧光信号的强弱来检测线粒体膜电位的变化和凋亡的发生,可用于培养的细胞或从组织中提取出的线粒体的膜电位检测。Tetramethylrhodamine, methyl ester (TMRM) 也是一种可透过细胞膜的阳离子荧光染料,单激光激发和单荧光发射峰。可用543nm激光激发,发射橙红色荧光波长在580nm左右。相对其他荧光探针,TMRM具有许多优点如染料在线粒体积累仅源于膜电位变化更;相对毒性更小;和细胞器结合率低;适合做线粒体膜电位的定量分析等。
缺血性脑卒中是人类死亡的主要原因之一,也是全球范围内成人致残的主要原因[1]。2019年我国有缺血性脑卒中患者2 418万例,给我国医疗卫生系统造成巨大负担[2]。缺血后大脑血液供应的中断会引发一系列病理生理改变。尽管恢复脑血流对挽救缺血组织至关重要,但血流恢复可能会进一步加重脑损伤[3]。再灌注损伤的机制包括活性氧(reactive oxygen species,ROS)的突然产生、自噬的激活和细胞因子的释放,其中线粒体功能障碍在介导这些病理生理过程中发挥重要作用[4]。目前,重组组织纤溶酶原激活剂(recombinant tissue plasminogen activator,rt-PA)已被批准应用于缺血性脑卒中的治疗[5]。由于治疗时间窗窄,能够应用rt-PA治疗的患者不足10%[6]。因此,寻找更有效的防治缺血性脑卒中的药物至关重要。 在中医理论中,缺血性脑卒中属于“中风”范畴。补阳还五汤是治疗缺血性脑卒中的经典方剂,已有数百年的临床应用历史[7-8]。方中以生黄芪为君药,补益元气,旨在气旺则血行,瘀去则络通,对中风之气虚血瘀证有显著疗效。现代药理学研究证实,芒柄花素是黄芪的主要活性成分之一,具有抗炎[9]、抗氧化[10]、神经保护[11]等作用。但是,水溶性差限制了其在中枢神经系统的应用。通过磺化反应合成的芒柄花素磺酸钠(sodium formononetin-3?-sulphonate,Sul-F)克服了其水溶性差的难题,为其应用于缺血性脑卒中的研究提供了物质基础。 本研究通过建立大脑中动脉栓塞模型(middle cerebral artery occlusion,MCAO)大鼠模型,考察Sul-F对大鼠脑缺血再灌注(ischemia-reperfusion, I/R)损伤的改善作用和对线粒体凋亡通路的影响,旨在探讨Sul-F是否通过调控线粒体凋亡通路改善脑I/R损伤。 1 材料 1.1 动物 SPF级雄性SD大鼠75只,体质量(300±10)g,购自斯贝福(北京)有限公司,许可证号SYXK(冀)2021-006。大鼠饲养于动物房,温度(25±1)℃,相对湿度(50±10)%,昼夜周期为12 h,自由摄食饮水,实验开展前适应性饲养1周。本实验所有操作均严格按照动物伦理要求进行,且经河北中医学院实验动物管理和伦理委员会批准(批准号DWLL202306001)。 1.2 药品与试剂 依达拉奉注射液(国药准字H20080056,批号2109050)购自国瑞药业有限公司;Sul-F(质量分数>95%,批号160901)由河北国金药业有限责任公司提供;2,3,5-氯化三苯基四氮唑(2,3,5-triphenyltetrazolium chloride,TTC)染液(批号1222A23)购自美国Sigma-Aldrich公司;苏木素-伊红(hematoxylin-eosin,HE)染液试剂盒(批号MD070823)购自碧云天生物技术有限公司;原位末端标记(TdT-mediated dUTP nick end labeling,TUNEL)检测试剂盒(批号061223240108)、线粒体膜电位(JC-1)检测试剂盒(批号C2006)购自碧云天生物技术有限公司;谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)测定试剂盒(批号A005-1)、超氧化物歧化酶(superoxide dismutase,SOD)测定试剂盒(批号A001-3)、丙二醛(malondialdehyde,MDA)测定试剂盒(批号A003-1)均购自南京建成生物工程研究所;电镜固定液(批号02607-BA)、包埋剂(批号90529-77-4)、醋酸双氧铀(批号H60602624A8)购自SPI公司;无水乙醇(批号100092183)、丙酮(批号10000418)购自国药集团化学试剂有限公司;铜网(批号BZ100205a)购自北京中镜科仪技术有限公司;锇酸(批号18456)购自Ted Pella公司;柠檬酸铅(批号180705-C5382)购自EMS公司;β-actin抗体(批号86e1489)、B淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)抗体(批号35y4418)、Bcl-2相关X蛋白(Bcl-2 associated X protein,Bax)抗体(批号43z8686)、半胱氨酸天冬氨酸蛋白酶-3(cystein-asparate protease-3,Caspase-3)抗体(批号53j2158)购自Affinity公司;HRP标记的山羊抗兔二抗(批号20220521)购自北京百奥思科生物科技有限公司;Trizol(批号342430AX)购自艾德莱公司;ExonScript First-Strand Synthesis SuperMix with dsDNase试剂盒(批号231106-A5)购自成都市蓉为基因生物科技有限公司;硅胶线栓(批号230701162)购自长沙迈越生物科技有限公司。 1.3 仪器 JT-12J型全自动脱水机、JB-L5型加热石蜡包埋系统(武汉俊杰电子有限公司);RM2235型石蜡切片机、UC7型超薄切片机(德国Leica公司);XS-2100型光学显微镜(NOVEL公司);ELCIPSE-CI型正置荧光显微镜(日本Nikon公司);7800型透射电子显微镜(日本Hitachi公司);LF-2000型SDS-PAGE电泳系统(北京龙方科技有限公司);JYO2S型凝胶成像系统(北京君意东方电泳设备有限公司);chemiscope 6100型化学发光成像系统(上海勤翔科学仪器有限公司);272005652型实时荧光定量[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]仪(美国Life Techologies公司);BeamCyte-1026型流式细胞仪(必达科生物科技有限公司);680型酶标仪(美国Bio-Rad公司)。 2 方法 2.1 动物造模、分组及给药 大鼠I/R损伤模型的建立参照文献方法[12],大鼠ip 1%戊巴比妥钠(50 mg/kg)麻醉后,置于加热垫上,保持肛温37 ℃。颈前皮肤备毛,在前正中线做切口,暴露颈总动脉、颈内动脉和颈外动脉,结扎颈外动脉远端,夹闭颈内动脉远端颈总动脉近端,将线栓经颈内动脉送入大脑中动脉,推送线栓18~20 mm,缺血2 h后拔出线栓并且结扎端口。再灌注0 h时,选择神经功能评分为1~3分的大鼠,随机分为模型组、依达拉奉(3 mg/kg)[13]组和Sul-F高、低剂量(80、40 mg/kg)组,每组15只。另取15只大鼠仅分离血管不插线栓作为假手术组。分别于再灌注0、12 h尾iv给药(4 mL/kg),假手术组和模型组给予等体积的生理盐水。再灌注24 h后取材进行后续实验。 2.2 神经功能评分 再灌注0、24 h时,采用盲法对各组大鼠进行Zea-Longa评分[14]:0分,活动基本正常;1分,提起时对侧前爪无法完全伸展;2分,向手术对侧转圈;3分,向手术对侧倾倒;4分,意识丧失。 2.3 TTC染色测定脑梗死体积 大鼠麻醉,断头取脑,?20 ℃冷冻30 min,以1.5~2 mm厚度进行冠状面切片。置于TTC染液玻璃皿中,37 ℃避光孵育30 min,吸出多余的TTC染液,倒入4%多聚甲醛固定过夜,相机拍照后用Image J软件进行分析,记录脑梗死面积(灰白色)及全脑面积(红色),计算脑梗死体积率[15]。 脑梗死体积率=(脑梗死面积×厚度)/(全脑面积×厚度) 2.4 HE染色观察脑组织病理变化 大鼠麻醉,断头取脑,4%多聚甲醛固定24 h,沉糖1周,石蜡包埋,以4 μm厚度行冠状切片,HE染色后于光学显微镜下进行观察与拍照。 2.5 TUNEL荧光染色观察脑组织细胞凋亡情况 将石蜡切片标本进行脱蜡、复水、抗原修复以及H2O2封闭,按照TUNEL凋亡检测试剂盒说明书进行TUNEL染色,并使用DAPI对细胞核进行复染。封片后置于荧光显微镜下观察与拍照,并计算细胞凋亡率。 细胞凋亡率=TUNEL阳性细胞数/DAPI阳性细胞数 2.6 JC-1探针检测脑缺血半暗带组织线粒体膜电位情况 取脑缺血半暗带组织,经300目钢网研磨、滤过到60 mm培养皿中,将滤过后的组织悬液转入15 mL新离心管中。加入Hank平衡盐溶液稀释至10 mL,反复吹打30次,冰上静置5 min,取上清液至15 mL新离心管中。滤过2次,滤液经1 000 r/min离心10 min,弃上清,加入Hank平衡盐溶液重悬,离心后弃上清。加入1 mL Hank平衡盐溶液重悬,细胞计数板计数,并调整细胞密度为1×106个/mL。取200 μL细胞,重悬于0.5 mL细胞培养液中。按照试剂说明书进行染色,流式细胞仪上机检测,以红绿荧光的比值表示线粒体膜电位变化。 2.7 透射电镜观察脑缺血半暗带组织超微结构 取缺血半暗带脑组织,剪成1 mm3小块,经PBS漂洗、4 ℃固定(2.5%戊二醛,12 h)、PBS漂洗、固定(1%锇酸,2 h)、PBS漂洗、丙酮脱水(30%、50%、70%、80%、95%、100%)、渗透包埋(Epon812)、超薄切片、染色(醋酸双氧铀和柠檬酸铅)后,在透射电子显微镜下采集图像分析。 2.8 ELISA检测脑缺血半暗带组织匀浆中MDA水平和SOD、GSH-Px活性 取缺血半暗带脑组织匀浆,按照试剂盒说明书进行操作,采用酶标仪测量吸光度(A)值,并计算MDA水平和SOD、GSH-Px活性。 2.9 qRT-[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]检测脑缺血半暗带组织Caspase-3、Bcl-2和Bax mRNA表达 取缺血半暗带脑组织,Trizol法提取总RNA,紫外分光光度计测定总RNA浓度。利用逆转录试剂盒Superscript III将RNA反转录成cDNA,加入引物,以β-actin为内参,荧光定量[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]仪扩增,采用2???Ct法计算缺血半暗带Caspase-3、Bcl-2和Bax mRNA表达水平。 图片 2.10 Western blotting检测脑缺血半暗带组织Caspase-3、Bcl-2和Bax蛋白表达 取缺血半暗带脑组织,加入裂解液提取蛋白,采用BCA试剂盒测定蛋白浓度。蛋白样品经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,转至PVDF膜,于5%牛血清白蛋白中封闭,分别加入Caspase-3、Bcl-2、Bax抗体(1∶1 000),4 ℃孵育过夜,洗膜后,加入二抗(1∶20 000),37 ℃孵育1 h。滴加ECL混合液反应4 min后,采用化学发光成像系统显影并对条带灰度值进行分析。 2.11 统计学分析 采用SPSS 20.0软件进行数据分析,正态分布计量资料以表示,非正态分布计量资料以M(P25~P75)表示。多组间均数比较,服从正态分布与方差齐者,采用单因素方差分析(One-way ANOVA),两两比较采用LSD检验;不服从正态分布者,采用Kruskal-Wallis检验。 3 结果 3.1 Sul-F对MCAO大鼠神经功能评分的影响 如表2所示,与假手术组比较,模型组大鼠神经功能评分显著升高(P<0.01);与模型组比较,依达拉奉组和Sul-F高剂量组大鼠神经功能评分均显著降低(P<0.01),Sul-F低剂量组神经功能评分无显著变化;依达拉奉组与Sul-F高剂量组比较,神经功能评分无统计学差异。 图片 3.2 Sul-F对MCAO大鼠脑梗死体积的影响 如图1-A、B所示,与假手术组比较,模型组大鼠脑梗死体积率显著升高(P<0.01);与模型组比较,依达拉奉组和Sul-F高剂量组大鼠的脑梗死体积率显著降低(P<0.05);依达拉奉组与Sul-F高剂量组比较差异无统计学意义。 图片 3.3 Sul-F对MCAO大鼠脑缺血半暗带病理损伤与细胞凋亡的影响 如图1-C所示,假手术组大鼠脑缺血半暗带神经细胞形态结构正常,结构致密,核仁清晰;模型组脑缺血半暗带神经细胞肿胀,空泡增多,细胞核大小不一,形态不规则,核固缩偏于细胞一侧,部分核溶解。与模型组比较,依达拉奉组与Sul-F高剂量组脑缺血半暗带组织病理损伤情况明显改善,Sul-F低剂量组脑缺血半暗带组织病理损伤情况无明显改善。 如图1-D、E所示,与假手术组比较,模型组脑缺血半暗带TUNEL阳性细胞数显著增多(P<0.01);与模型组比较,各给药组TUNEL阳性细胞数显著降低(P<0.01);依达拉奉组与Sul-F高剂量组比较,TUNEL阳性细胞数无统计学差异。 3.4 Sul-F对MCAO大鼠脑缺血半暗带线粒体膜电位变化的影响 如图2所示,与假手术组比较,模型组大鼠脑缺血半暗带线粒体膜电位显著降低(P<0.01);与模型组比较,依达拉奉组和Sul-F高剂量组线粒体膜电位显著升高(P<0.05),两组之间比较无明显差异。 图片 3.5 Sul-F对MCAO大鼠脑组织超微结构的影响 如图3所示,假手术组大鼠脑组织线粒体丰富,分布均匀,线粒体嵴清晰可见,未见明显损伤的线粒体,无典型的线粒体自噬结构。模型组线粒体总体数量减少,线粒体嵴不清晰、水肿,可见典型自噬小体形成。与模型组比较,依达拉奉组线粒体数量增多,大部分线粒体嵴清晰,偶见线粒体自噬结构;Sul-F低剂量组线粒体数量增多,一部分线粒体嵴清晰,另一部分线粒体水肿,可见典型自噬小体形成;Sul-F高剂量组线粒体数量增多,大部分线粒体嵴清晰,偶见线粒体自噬结构。 图片 3.6 Sul-F对MCAO大鼠脑缺血半暗带组织氧化应激水平的影响 如图4所示,与假手术组比较,模型组大鼠脑缺血半暗带区组织MDA水平显著升高(P<0.01),SOD和GSH-Px活性显著降低(P<0.01);与模型组比较,各给药组MDA水平显著降低(P<0.05、0.01),SOD活性显著升高(P<0.01);Sul-F高剂量组GSH-Px活性显著升高(P<0.01)。 图片 3.7 Sul-F对MCAO大鼠脑缺血半暗带组织Bcl-2、Bax和Caspase-3 mRNA和蛋白表达的影响 如图5所示,与假手术组比较,模型组大鼠缺血半暗带区Caspase-3和Bax mRNA表达水平显著升高(P<0.01),Bcl-2 mRNA表达水平显著降低(P<0.01);与模型组比较,各给药组大鼠缺血半暗带区Caspase-3和Bax mRNA表达水平显著降低(P<0.01),Bcl-2 mRNA表达水平显著升高(P<0.01);与依达拉奉组比较,Sul-F高剂量组大鼠缺血半暗带区Caspase-3、Bax和Bcl-2 mRNA表达水平无显著差异。 图片 如图6所示,与假手术组比较,模型组大鼠缺血半暗带区Caspase-3和Bax蛋白表达水平显著升高(P<0.01),Bcl-2蛋白表达水平显著降低(P<0.01);与模型组比较,各给药组大鼠缺血半暗带区Caspase-3和Bax蛋白表达水平显著降低(P<0.01),Bcl-2蛋白表达水平显著升高(P<0.01);与依达拉奉组比较,Sul-F高剂量组大鼠缺血半暗带区Caspase-3、Bax和Bcl-2蛋白表达水平无显著差异。 图片 4 讨论 缺血性脑卒中占所有脑卒中的80%~85%,具有较高的致死率和致残率[16]。目前,缺血性脑卒中的治疗手段有机械取栓和药物溶栓,以尽快恢复脑血流、挽救缺血脑组织,但复流复氧可能加重脑组织损伤[3,17]。脑I/R损伤会引发一系列的病理反应,如离子失衡、细胞膜通透性改变、能量代谢障碍等,最终触发凋亡程序,引发细胞凋亡。细胞凋亡涉及线粒体通路和死亡受体通路,其中线粒体通路是启动凋亡程序的关键。因此,改善线粒体功能、保护神经元是治疗脑I/R损伤的关键。 正常情况下,线粒体产生的自由基及其清除处于动态平衡,在缺血等应激条件下,ROS生成过多,氧化还原平衡受损线粒体膜脂质过氧化和结构破坏,造成线粒体功能紊乱和氧化应激损伤[17]。SOD与GSH-Px是机体抗氧化酶系统的重要物质,通过清除线粒体ROS来减轻氧化应激损伤。线粒体膜电位(mitochondrial membrane potential,MMP)是由三羧酸循环产生的能量传递给电子并经呼吸链传递过程中,将质子从线粒体内膜的基质侧泵到内膜外所形成的跨膜电位差,MMP的下降是线粒体氧化应激损伤的早期指标[18]。本研究显示,在脑I/R损伤时,脑组织线粒体受损,主要表现有线粒体肿胀变形、线粒体嵴结构不清,可见线粒体自噬小体。同时,线粒体膜电位显著下降,氧化应激反应被激活,MDA含量显著升高,SOD和GSH-Px活力显著降低。 线粒体途径作为细胞凋亡的关键途径,主要受到Bcl-2家族和Caspase家族相关基因的调控。线粒体功能受损产生大量ROS,ROS过度累积会触发线粒体膜通透性转换孔打开,线粒体膜电位下降[19]。Bcl-2家族蛋白通过调节线粒体外膜通透性在细胞内凋亡信号转导中发挥重要作用。Bax和Bcl-2是Bcl-2家族的主要成员,二者通常以异源二聚体的形式存在。MMP升高时,Bcl-2表达上调,抑制细胞色素C释放以维持线粒体平衡,当Bax高表达时,MMP降低,线粒体中的细胞色素C释放入胞质中,在三磷酸腺苷(adenosine triphosphate,ATP)和dATP的协助下生成凋亡复合物,招募并启动Caspase-9,Caspase-9解体形成cleaved Caspase-9并进一步启动Caspase-3,激活下游Caspase级联瀑布,启动线粒体介导的细胞凋亡[20-21]。Caspase-3是Caspase家族中参与细胞凋亡的关键酶,可导致线粒体膜通透性增加、DNA断裂和染色质浓缩,可能是缺血性神经元核降解的关键执行者[17]。本研究中,脑I/R损伤使促凋亡基因Bax表达显著上调,抑凋亡基因Bcl-2表达显著下调,Caspase-3表达显著上调,提示脑I/R损伤时,线粒体介导的细胞凋亡途径启动。 芒柄花素是经典名方补阳还五汤君药黄芪的活性成分之一。体外研究发现,芒柄花素能够抑制多腺苷二磷酸核糖聚合酶1/凋亡诱导因子/蛋白激酶B(poly-adenosine diphosphate ribose polymerase/ apoptosis inducing factor/protein kinase B,PARP1/ AIF/Akt)信号通路减轻糖氧剥夺/复氧复糖条件下HT22小鼠神经元细胞损伤[22]。为提高其水溶性和生物利用度,经磺化反应合成芒柄花素磺酸钠。前期研究证实,Sul-F能够通过血脑屏障、低毒[23-25]。本研究发现,Sul-F能够减低脑I/R损伤大鼠的神经功能评分、降低脑梗死体积、减轻脑缺血半暗带的病理损伤及细胞凋亡,进而改善脑I/R损伤。 线粒体功能障碍是脑缺血再灌注诱导神经元死亡的标志之一[26],因此机制研究旨在探讨Sul-F对线粒体介导的细胞凋亡途径的影响。结果显示,Sul-F能够降低脑组织氧化应激水平,部分逆转线粒体膜电位的降低,改善线粒体超微结构损伤,调节Bcl-2/Bax平衡,降低Caspase-3表达,进而抑制线粒体凋亡途径,降低细胞凋亡,对脑I/R损伤具有潜在的治疗价值。 本研究以大鼠大脑中动脉栓塞模型模拟脑I/R损伤,结果显示Sul-F干预能够降低脑组织氧化应激水平,部分逆转线粒体膜电位的降低,改善线粒体超微结构损伤,降低细胞凋亡,改善脑梗死体积和神经功能评分。进一步研究发现,Sul-F能够降低Bax表达、升高Bcl-2表达,降低下游Caspase-3表达,抑制线粒体凋亡信号通路。综上,Sul-F通过调控Bcl-2/Bax平衡,降低线粒体介导的细胞凋亡,改善脑缺血再灌注损伤。
以下问题由版友liugw123450提供:“有没有人会用到zeta电位仪,显微镜电泳法的与流动电势法的固体薄膜电位仪”见:主题帖[color=#DC143C]“应用现代激光衍射技术的粒径测量方法”5楼[/color]为鼓励liugw123450版友,转送2个积分。
除膜片钳外,能够测定膜电位的仪器及方法有?
乳腺癌是世界范围内女性最常见的致死性恶性肿瘤,据统计,2020年女性乳腺癌已超越肺癌成为全球癌症发病率最高的癌种[1-2]。其中三阴性乳腺癌(triple-negative breast cancer,TNBC)是雌激素受体(estrogen receptor,ER)、孕激素受体(progesterone receptor,PR)和人表皮生长因子受体2(human epidermal growth factor receptor 2,HER-2)均呈阴性表达的乳腺癌亚型,占所有乳腺癌的15%~20%[3],具有侵袭力强、转移率高、术后复发率高、预后差的特点[4]。由于TNBC内分泌治疗的不确定性及靶向治疗的不应答性,导致临床上的治疗效果不理想[5-6]。因此,寻找有效抑制TNBC增殖转移的药物、降低患者的病死率一直是乳腺癌基础研究的一个重要方向[7-8]。 石蒜碱是石蒜Lycoris radiata (L'Hér.) Herb.、文殊兰Crinum asiaticum L. var. sinicum (Roxb. et Herb.) Baker、朱顶红Hippeastrum rutilum (Ker.-Gawl.) Herb.等石蒜属植物鳞茎中含量较高的异喹啉类生物碱,具有抗肿瘤、抗病毒、抗菌、抗炎镇痛、保肝等药理活性[9-10],近年来石蒜碱的抗肿瘤作用受到众多研究者的关注。有文献报道石蒜碱对人乳腺癌MCF-7细胞[11]、人宫颈癌Hela细胞[12-13]、人肝癌HepG-2细胞[13-16]、人胃癌SGC-7901细胞[17]、人结肠腺癌LoVo细胞[18-19]具有显著的抑制作用,但对其作用机制的研究仍然处于初始阶段。本研究以人乳腺癌MDA-MB-231细胞为研究对象,主要通过体外实验从细胞水平和分子水平探讨石蒜碱对MDA-MB-231细胞的体外抑制活性及其通过线粒体氧化损伤诱导肿瘤细胞自噬及凋亡的机制,为今后石蒜碱抗肿瘤新药的深入研发和临床实践提供理论基础和实验参考。 1 材料 1.1 细胞株 MDA-MB-231细胞由国家教育部抗肿瘤天然药物工程技术研究中心提供。 1.2 药品与试剂 石蒜碱(批号34296,质量分数98%)购自阿拉丁试剂有限公司;胎牛血清(批号0201021)购自浙江杭天生物科技公司;RPMI 1640细胞培养基(批号AD123707271)购自美国HyClone公司;二甲基亚砜(dimethyl sulfoxide,DMSO,批号20200901)购自天津中和盛泰化工有限公司;Hoechst 33258染液(批号C1011)、SDS-PAGE蛋白上样缓冲液(批号P0015)、吉姆萨染液(批号C0131)、CCK-8试剂盒(批号C0043)、活性氧(reactive oxygen species,ROS)检测试剂盒(批号S0033S)、PMSF(批号ST505)、HRP标记的山羊抗大鼠IgG二抗(批号A0192)、Western blotting及IP细胞裂解液(批号072318180723)、30% Acr-Bis(批号093018181017)购自碧云天生物技术研究所;碘化丙啶(propidium iodide,PI)染液(批号R20285)、Rhodamine 123(批号R8004)购自美国Sigma公司;台盼蓝(批号72-52-1)购自美国默克公司;Reagent A染液(批号5000113)购自北京诺博莱德科技有限公司;聚山梨酯20(批号20190207)购自美国Biotopped公司;Tris(批号181127)购自美国Amresco公司;兔抗半胱氨酸天冬氨酸蛋白酶-3(cystein-asparate protease-3,Caspase-3)抗体(批号WL02512)、兔抗B淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)抗体(批号WL01506)、兔抗Bcl-2相关X蛋白(Bcl-2 associated X protein,Bax)抗体(批号WL02385)、兔抗细胞色素C(cytochrome-C,Cyt-C)抗体(批号WL04963)、兔抗甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体(批号WL01114)购自沈阳万类生物科技有限公司;兔抗线粒体内膜转位酶(translocase of inner membrane,TIM)抗体(批号PSI-RF16109)、兔抗线粒体外膜转位酶(translocase of outer membrane,TOM)抗体(批号PSI57577)、兔抗E3泛素连接酶(E3 ubiquitin protein ligase,PARKIN)抗体(批号PSI50248)、兔抗PTEN诱导的激酶1(PTEN induced putative kinase 1,PINK1)抗体(批号PSI7859)、兔抗微管相关蛋白轻链3(microtubule-associated protein light chain 3,LC3-B)抗体(批号BS79705)、兔抗p62抗体(批号p196-269)购自艾美捷科技有限公司。 1.3 仪器 ECO-170P-230型细胞培养箱、Model 680型酶标仪(美国NBS公司);Adventurer型万分之一电子天平(美国OHAUS公司);EPICS-XL型流式细胞仪、AllegraTM 64R型低温高速离心机(美国Beckman-Coulter公司);CKX-41-32型倒置显微镜(日本Olympus公司);荧光显微镜、TCS-SP2激光共聚焦扫描显微镜(德国Leica公司);680型全自动酶标仪(美国Bio-Rad公司);P型微量[url=https://insevent.instrument.com.cn/t/9p][color=#3333ff]移液器[/color][/url](芬兰百得公司);标准型PB-10 pH计(德国Sartorius公司);GIS-2019型Tannon凝胶成像系统(天能科技有限公司);DYY-7C型电泳仪、M344039型垂直电泳转印槽(北京六一仪器厂)。 2 方法 2.1 细胞培养 MDA-MB-231细胞复苏后接种于含10%胎牛血清的RPMI 1640培养基中,置于5% CO2、37 ℃恒温培养箱中培养,待细胞长势良好时进行传代,取对数生长期的细胞进行实验。 2.2 CCK-8法检测细胞增殖活性 MDA-MB-231细胞以2×103个/孔接种于96孔板中,细胞培养24 h后,给药组每孔加入不同浓度(2、4、8、16、32 μmol/L)的石蒜碱100 μL,对照组加入100 μL细胞培养基,每组均设置6个平行孔,处理48 h后,每孔加入10 μL CCK-8试剂,继续培养4 h。采用酶标仪检测490 nm处的吸光度(A)值,计算各组细胞的增殖抑制率与石蒜碱对MDA-MB-231细胞的半数抑制浓度(half inhibitory concentration,IC50)。 2.3 倒置显微镜、荧光显微镜、激光共聚焦扫描显微镜观察细胞形态变化 MDA-MB-231细胞以3×103个/孔分别接种于2块6孔板中,细胞培养24 h后,根据石蒜碱对MDA-MB-231细胞的IC50设定3个给药剂量,分别以3、6、12 μmol/L的给药浓度每孔加入石蒜碱1 mL,对照组加入1 mL细胞培养基,继续处理48 h。取1块板用倒置显微镜观察并拍照后,每孔加入1 mL多聚甲醛固定1 h,冲洗后加入200 μL Hoechst 33258染液,37 ℃孵育30 min后,用荧光显微镜观察并拍照;取另1块板收集各组细胞,用预冷的PBS重悬细胞并弃去上清液,加入Annexin V-FITC于37 ℃避光孵育15 min,冲洗后加入PI染液于4 ℃避光孵育15 min后,用激光共聚焦扫描显微镜观察并拍照。 2.4 集落实验检测细胞克隆能力 MDA-MB-231细胞以1×103个/孔接种于6孔板中,细胞培养24 h后,按“2.3”项下方法对细胞进行分组和给药,连续培养7 d后弃去培养基。PBS洗涤后用甲醇固定10 min,冲洗后加入吉姆萨染液染色后,用倒置显微镜观察细胞集落形成率并拍照。 2.5 划痕实验检测细胞迁移能力 MDA-MB-231细胞以1×105个/孔接种于6孔板中,细胞培养24 h,细胞融合至70%~80%后,用200 μL[url=https://insevent.instrument.com.cn/t/9p][color=#3333ff]移液器[/color][/url]倚靠直尺,枪头垂直于每孔底部竖直划痕。PBS冲洗后,按“2.3”项下方法对细胞进行分组和给药,培养48 h后,用倒置显微镜观察细胞的迁移情况并拍照记录,比较各组间的划痕宽度,使用Image J软件测量并计算划痕愈合率。 2.6 流式细胞仪检测细胞凋亡率 按“2.3”项下方法对细胞进行分组和给药,培养48 h后,收集各组细胞,加入70%冷乙醇2 mL于4 ℃固定24 h后离心。弃去上清液,PBS冲洗后,加入800 μL PI染液,4 ℃避光孵育30 min,经尼龙网滤过后,采用流式细胞仪进行检测,激发波长为488 nm。 2.7 流式细胞仪检测ROS水平 按“2.3”项下方法对细胞进行分组和给药,培养48 h,收集各组细胞,PBS洗涤后加入5 μmol/L DCFH-DA染液0.2 mL,37 ℃避光孵育20 min,经尼龙网滤过后,采用流式细胞仪进行检测。 2.8 流式细胞仪检测线粒体膜电位 按“2.3”项下方法对细胞进行分组和给药,培养48 h后,收集各组细胞,PBS洗涤后,避光加入Rhodamine 123染料,避光孵育30 min后离心弃去上清液,用PBS洗涤并混匀细胞,经尼龙网滤过后,采用流式细胞仪进行检测。 2.9 激光共聚焦扫描显微镜检测线粒体膜通透性转换孔(mitochondrial permeability transition pore,MPTP)活性 按“2.3”项下方法对细胞进行分组和给药,培养48 h后,收集各组细胞,加入37 ℃预热的Reagent A染液500 μL,离心后弃去上清液。37 ℃避光加入染色工作液,混匀后孵育20 min,离心去除上清液,将细胞吹打混匀后,经尼龙网滤过,采用激光扫描共聚焦显微镜检测并进行拍照。 2.10 Western blotting检测线粒体自噬相关蛋白TIM、TOM、PARKIN、PINK1、LC3-B、p62和凋亡相关蛋白Caspase-3、Bax、Bcl-2、Cyt-C表达 按“2.3”项下方法对细胞进行分组和给药,培养48 h后,收集各组细胞,加入含PMSF的细胞裂解液,冰上裂解30 min后将细胞加入EP管中,离心15 min。取上清液,煮沸使蛋白变性,采用BCA试剂盒定量蛋白浓度。采用SDS-PAGE凝胶电泳,转至PVDF膜,5%脱脂奶粉封闭2 h后,加入一抗,4 ℃孵育过夜。TBST洗膜后加入二抗,37 ℃孵化2 h,洗膜后加入化学发光试剂,采用凝胶成像系统拍照并进行分析。 2.11 统计学分析 用SPSS 21.0软件进行统计分析,数据以表示,多样本均数比较采用One-way ANOVA分析,通过Graphpad Prism 8软件绘图。 3 结果 3.1 石蒜碱对MDA-MB-231细胞增殖的影响 如图1所示,石蒜碱对MDA-MB-231细胞具有显著的增殖抑制作用(P<0.01),且呈浓度相关性。石蒜碱对MDA-MB-231细胞的IC50为6.21 μmol/L,并参考IC50值设定后续石蒜碱给药浓度分别为3、6、12 μmol/L。 3.2 石蒜碱对MDA-MB-231细胞形态的影响 采用倒置显微镜、荧光显微镜、激光共聚焦显微镜观察石蒜碱对MDA-MB-231细胞形态的影响,如图2所示,与对照组比较,石蒜碱给药后,随着给药浓度增加,细胞生长逐渐变稀疏,细胞膜破裂现象更加明显,细胞间轮廓更加模糊,细胞核固缩形成凋亡小体,发出较强荧光。 3.3石蒜碱对MDA-MB-231细胞克隆、迁移的影响 集落实验结果表明,石蒜碱可以抑制MDA-MB-231细胞的克隆能力(图3-A),且随着浓度的增加细胞集落数量逐渐减少,且呈浓度相关性。划痕实验结果显示,石蒜碱可以显著抑制MDA-MB-231细胞的迁移能力(P<0.01,图3-B、C),呈剂量相关性。 3.4 石蒜碱对MDA-MB-231细胞凋亡率、ROS水平的影响 如图4-A、B所示,经流式细胞仪PI单染法检测出现明显的凋亡峰,表明DNA的合成受到抑制,且随着给药浓度增加,凋亡峰越明显,凋亡率也呈上升趋势,与对照组比较有显著性差异(P<0.01),且呈浓度相关性。如图4-C、D所示,随着给药浓度增加,细胞内ROS水平逐渐升高,具有显著性差异(P<0.01),且呈浓度相关性。 3.5 石蒜碱对MDA-MB-231细胞线粒体膜电位和MPTP的影响 如图5-A、B所示,经流式细胞仪检测,随着石蒜碱给药浓度增加,细胞内线粒体膜阳性表达率逐渐降低,具有显著性差异(P<0.01),且呈浓度相关性。如图5-C、D所示,应用激光扫描共聚焦显微镜结合AM染色技术对不同浓度的石蒜碱作用48 h后的MDA-MB-23细胞进行检测,激光扫描共聚焦显微镜扫描得到的荧光象素强度反映出细胞膜通透性的改变,随着给药浓度增加,细胞内线粒体膜通透性逐渐升高,具有显著性差异(P<0.01),且呈浓度相关性。 3.6 石蒜碱对MDA-MB-231线粒体自噬和凋亡相关蛋白表达的影响 应用凝胶成像系统分析MDA-MB-231细胞中线粒体自噬和凋亡相关蛋白表达的情况。如图6所示,随着石蒜碱浓度增加,细胞自噬相关蛋白TIM、TOM和p62蛋白表达量逐渐降低,PARKIN、PINK1和LC3-B蛋白表达量逐渐升高,均具有显著性差异(P<0.01)。如图7所示,随着石蒜碱浓度增加,细胞凋亡相关蛋白Bcl-2蛋白表达量逐渐降低,Bax、Caspase-3和Cyt-C蛋白表达量逐渐升高,均具有显著性差异(P<0.01)。 4 讨论 乳腺癌已成为全球最常见的恶性肿瘤,与乳腺癌的其他分子亚型相比,TNBC最具侵袭性和高度异质性[20-22],使其在临床上难以得到有效治疗。因此如何有效抑制TNBC侵袭、增殖和转移是目前亟待解决的问题。近年来,有研究表明中药在抗肿瘤方面具有显著的优势[23-25]。石蒜碱是异喹啉类生物碱,广泛分布于石蒜属植物鳞茎中,具有较强的抗肿瘤活性[26-27]。基于石蒜碱的抗肿瘤作用,结合课题组前期研究中TNBC细胞活性筛选,发现石蒜碱对MDA-MB-231细胞较为敏感,故选择MDA-MB-231细胞作为研究对象,本研究结果发现石蒜碱对MDA-MB-231细胞的增殖和迁移具有显著抑制作用,且呈浓度相关性。 ROS水平升高和线粒体功能障碍是诱导肿瘤细胞自噬和凋亡的重要途径[28]。研究发现,过量ROS的产生会诱发肿瘤细胞的损伤、自噬及凋亡并降低细胞的多药耐药性[29]。此外,肿瘤细胞对外源性ROS比正常细胞更敏感且ROS具有一定的细胞毒性。因此,促进ROS水平升高的药物可表现出一定的抗癌活性。有研究表明,线粒体功能障碍与多种恶性肿瘤的发生及ROS的过量产生密切相关[30]。本研究通过激光共聚焦显微镜和流式细胞仪检测结果表明,石蒜碱可以显著提高MDA-MB-231细胞凋亡率和ROS水平,并使线粒体膜电位下降,MPTP开放。这表明石蒜碱诱导细胞自噬和凋亡作用可能与线粒体的氧化损伤有关。 TOM及TIM是线粒体膜蛋白,当线粒体自噬增强时,其细胞内表达水平下降。研究表明线粒体损伤会使线粒体膜电位降低,导致PINK1在线粒体外膜上表达,从而使PINK1-PARKIN依赖性线粒体自噬反应被激活[31]。LC3-B是自噬体形成的特异性标志物,其含量与自噬泡数量成正比,因此被广泛用于监测细胞自噬。p62作为自噬降解的产物,自噬增强,p62水平会下降。p62还可与自噬体膜上的LC3-B蛋白及泛素化的蛋白形成复合物,在自噬溶酶体内完成降解[32]。ROS的过度累积,会触发MPTP开放,导致线粒体膜电位下降,引起Cyt-C从线粒体释放并进入细胞质中,进而激发Caspase的级联反应并启动细胞线粒体凋亡[33]。Bcl-2为抗凋亡蛋白,Bax为促凋亡蛋白,当接收到凋亡刺激信号后可转位至线粒体膜上,Bcl-2和Bax可形成二聚体或多聚体,从而增加细胞线粒体膜的通透性,进一步激活Caspase级联反应,Caspase-3可通过抑制凋亡抑制物,从而破坏细胞结构使蛋白丧失功能[34]。本研究通过Western blotting检测自噬相关蛋白和凋亡相关蛋白的表达,结果显示石蒜碱能够上调PARKIN、PINK1、LC3-B、Caspase-3、Bax和Cyt-C蛋白表达,下调TIM、TOM、p62和Bcl-2蛋白表达,表明石蒜碱可通过线粒体的氧化损伤介导MDA-MB-231细胞的自噬及凋亡。 综上,石蒜碱对MDA-MB-231细胞具有生长抑制作用,并可通过调控线粒体氧化损伤介导MDA-MB-231细胞的自噬及凋亡。本研究为石蒜碱抗肿瘤新药的深入研发和临床实践提供理论基础。
[b][size=15px][color=#595959]急性肝损伤(ALI)[/color][/size][/b][size=15px][color=#595959]是突然和大量肝细胞损伤的严重后果。线粒体是多种药物和化学物质引起的药物性肝毒性的重要靶点。最近的证据表明,[b]线粒体功能[/b]异常可触发各种肝脏疾病的发生,并有助于感染、毒素和药物滥用引起的ALI 。线粒体扰动也会影响肝功能的恢复,改善线粒体功能一直是探索包括ALI在内的各种肝脏疾病潜在治疗方法的热门研究领域。[/color][/size] [b][size=15px][color=#595959]金钗石斛(DNL)[/color][/size][/b][size=15px][color=#595959]是载入《中国药典》(2020年版)的著名中药。先前的数据表明,[b]金钗石斛生物碱(DNLA)[/b]通过减少[b]氧化应激[/b]和改善线粒体功能来保护CCl4诱导的肝损伤,但确切的调控信号通路尚不清楚。[/color][/size][size=15px][color=#595959][/color][/size] [size=15px][color=#595959]该研究旨在探讨[b]坏死性凋亡[/b]在CCl4诱导的肝损伤模式中的作用,并确定DNLA是否通过抑制线粒体ROS (mROS)介导的坏死性凋亡对CCl4诱导的急性肝损伤(ALI)有保护作用。 [/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959]从DNL中提取DNLA,采用[url=https://insevent.instrument.com.cn/t/5p][color=#3333ff]液相色谱[/color][/url]-质谱([url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url])法测定其含量。采用C57BL/6J小鼠进行体内实验。先给药DNLA (20 mg/kg/d, ig) 7 d,然后给药CCl4(20 μL/kg, ip)。通过小鼠血清生化指标评估和苏木精伊红(H&E)染色对肝组织进行组织病理学检查,评价CCl4对小鼠肝损伤的影响。采用western blotting和实时荧光定量[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url] (RT-q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url])检测蛋白和基因表达。使用荧光探针DCFH-DA检测活性氧(ROS)产生,使用荧光探针JC-1评估线粒体膜电位。采用荧光探针MitoSOX检测[b]线粒体ROS (mROS)水平[/b]。 [/color][/size] [b][size=15px][color=#595959][/color][/size][/b][size=15px][color=#595959]DNLA减轻CCl4诱导的肝损伤,表现为降低AST和ALT水平,改善肝脏病理。[b]DNLA通过降低RIPK1、RIPK3和MLKL磷酸化抑制坏死性凋亡,同时增强线粒体功能[/b]。它还打破了mROS与RIPK1/RIPK3/MLKL激活之间的正反馈循环。在白藜芦醇和线粒体SOD2过表达中观察到类似的结果,两者都减轻了mROS和坏死性凋亡。进一步的机制研究发现,DNLA抑制RIPK1的氧化并降低其磷酸化水平,从而降低RIPK3和MLKL的磷酸化水平,阻断坏死性凋亡,减轻肝损伤。[/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size] [b][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][/b][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][size=16px][/size][/color][/size][b][size=15px][color=#595959][/color][/size][size=15px][color=#595959]DNLA通过减少mROS介导的RIPK1氧化,从而降低RIPK1、RIPK3和MLKL的磷酸化,抑制坏死性凋亡信号通路,对肝损伤具有保护作用[/color][/size][/b][size=15px][color=#595959]。[/color][/size][size=15px][color=#595959][/color][/size]
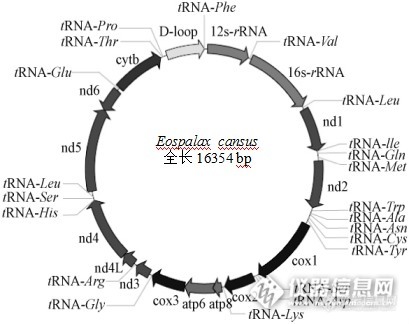
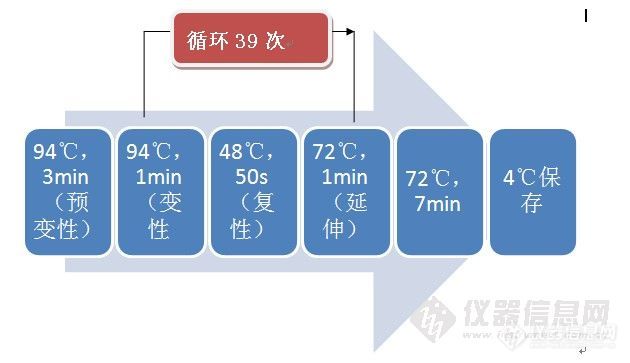
[b][/b][align=center]鼢鼠([i]Eospalax[/i])线粒体基因组测定及注释分析[/align][align=center]西安国联质量检测技术股份有限公司[/align][align=center]安平中心:李瑞[/align][b]摘要【[/b]目的】获得鼢鼠线粒体基因组全序列,为线粒体基因组功能标记及进化生物学等研究提供基础资料。【方法】参考鼹型鼠等动物的线粒体基因组序列,设计出可覆盖鼢鼠线粒体基因组的16对引物,采用[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]产物直接测序法测得甘肃鼢鼠线粒体基因组全序列,分析其基因组的特点和基因结构。并结合GenBank中发表的啮齿类动物基因组全序列,探讨啮齿类动物的系统进化关系。【结果】鼢鼠线粒体基因组全长16354bp,其中包括22个tRNA基因、13个蛋白质编码基因、2个rRNA基因和2个D-loop区。碱基组成为33.5%A、24.2 %C、12.3 %G、30.0 %T。【结论】鼢鼠线粒体基因组结构及其信息和其他啮齿类动物的结构一致,线粒体变异保守。研究结果为鼢鼠的低氧适应、系统发育关系等提供了基础资料。[b]关键词 [/b]鼢鼠;线粒体基因组;序列分析 鼢鼠([i]Eospalax[/i])是分布于我国的主要啮齿类动物之一,其体型较小,栖息于洞穴内有挖掘活动,扩散能力强,数量波动大,是生态系统中重要的初级消费者,处于生态系统中的中心位置,草原生态系统中其能流比重很大[sup][/sup]。动物线粒体([color=#333333]Mitochondrion[/color])基因组为双链闭合环状分子[sup][/sup],少数也有线性的,它们具有分子量相对较小、结构简单、缺少重组、母性遗传和进化速率快等特点,已成为动物系统发育与进化、群体遗传学、分子生态学以及疾病机理研究等领域的理想材料[sup][/sup]。甘肃鼢鼠是仅分布于我国西北部的土著物种,其外形似中华鼢鼠,主要分布于甘肃临潭县及其附近地区。目前对线粒体DNA的研究主要在动物分子遗传学、分子生态学、种群遗传结构分析、遗传多样性、物种和品系鉴定、保护遗传学等方面得到了广泛应用[sup][4[/sup][sup],[/sup][sup]5][/sup]1. [b]实验材料和方法[/b]1.1 实验材料鼢鼠:采集于天祝(经度102.84、纬度 37.2)1个群体;鼢鼠解剖采集肝脏及肌肉组织样品,-20℃保存备用。1.2 线粒体DNA的提取用剪刀将肝脏及肌肉材料剪成小块,取0.1cm左右的小块肝脏及肌肉材料,采用常规的SDS/蛋白酶K裂解,酚氯仿提取DNA[sup][/sup],使用琼脂糖凝胶电泳检测其完整性。1.3 引物设计和[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]扩增通过Clustal X1.83比对,寻找相对应保守区域位置,用Primer Premier5.0引物设计软件设计引物,并对每条引物进行评价和修改,最终确定16对引物。以所提取的DNA为模板,用16对引物扩增覆盖整个线粒体基因组。利用引物进行[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]扩增,反应体系总体积为50μL,其中含有6μL [url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url] buffer、3μL MgCl[sub]2[/sub](1.5mmol)、MgCl[sub]2[/sub],2μL dNTPs (100μL mol)、上下游引物各2μL (0.25μL mol)、Taq DNA聚合酶2μL (1U)、总DNA约为2μL (25ng)、去离子水31μL。反应程序为:94℃预变性4 min,94℃变性50s,48-45℃退1min,72℃延伸1 min 30s,循环30次,之后72℃延伸10min,并根据不同引物的退火温度和扩增反应的实际效果进行优化。取 5 μL [url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]扩增产物,和2 μL DNA marker 2000,进行1.0%琼脂糖凝胶(1×TBE)5V/电泳,用紫外观察[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]产物扩增情况,凝胶成像仪扫描记录结果。1.4 纯化、测序和序列拼接 在[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]产物中加入5 U SAP和2 U ExoⅠ,震荡混匀,37℃保温1 h,然后75℃保温15 min以灭活SAP和ExoⅠ酶,纯化好的模板可以在4℃保存24 h或-20℃长期保存。将纯化后的引物送往上海生工生物技术服务有限公司用ABI-3730序列自动分析仪进行双向测序。利用DNASTAR和测序峰图结果分析软件Chromas 2.22校对测序图,DNAMAN拼接序列。得到甘肃鼢鼠线粒体全基因组全序列。2. [b]结果[/b]2.1 鼢鼠线粒体基因组基因定位2.2.1 鼢鼠线粒体2个rRNA的分析哺乳动物线粒体的rRNA具有高度的保守性,它们的位置固定,12S rRNA位于tRNA-phe 和tRNA-Val之间,16S rRNA位于tRNA-Val和 tRNA-Leu之间,12S rRNA起始位置为68,终止位置为1019,长度为952bp,16S rRNA起始位置为1086,终止位置为2651,长度为1566。同时我们比对了鼢鼠和中华鼢鼠的rRNA基因和蛋白质基因,12S rRNA和16S rRNA的相似性分别为91.0%和87.3%,高于蛋白质编码基因之间的相似性。2.2.3 鼢鼠线粒体基因组结构 除NADH脱氢酶亚基6外均在H链上,虽然鼢鼠染色体数目少、染色体大,但与其它哺乳动物线粒体全基因组相比,它的线粒体基因组的结构与其它哺乳动物是十分相似的。甘肃鼢鼠线粒体基因组结构见图1。[align=center][img=,409,324]http://ng1.17img.cn/bbsfiles/images/2017/09/201709081454_02_2904018_3.png[/img][/align]注:ND: NADH脱氢酶亚基(NADH dehydrogenase subunit)、Cox:细胞色素氧化酶亚基(cytochrome oxidase subunit)、Atp:ATP合成酶亚基(ATP synthase F0 subunit)、Cyt b:1个细胞色素b编码基因(cytochrome b)。[align=center][b]图1[/b] 甘肃鼢鼠线粒体基因组结构简图[/align][align=center]Fig.1 The gene organization of [i]Eospalax cansus[/i] mitochondrial genome[/align]3. [b] 讨论[/b] 甘肃鼢鼠线粒体基因组的D-loop区,长度为933bp,比中国地鼠D-loop区(867bp)长。D-loop区对目的基因是不可缺少的,虽然D-loop区不能编码蛋白质但对于遗传信息表达是不可缺少的,在它上面有调控遗传信息表达的核苷酸序列,具有遗传效应的,比如RNA聚合酶结合位点是具有遗传效应的。8只甘肃鼢鼠中有5个单倍型:3只临潭群体共享1个单倍型,2只天祝群体独享单倍型;其余个体均独享单倍型,表明了甘肃鼢鼠线粒体DNA D-loop区碱基变异快、进化快的特性,符合啮齿动物线粒体变异大的现象。随着研究的深入,以线粒体DNA中完整的基因序列或多个基因序列协同而获得遗传信息来探讨物种的系统进化关系,将是以后研究发展的主要方向[sup][/sup]。目前,线粒体DNA已经在许多哺乳类动物的起源进化的研究中取的了重大进展,而对甘肃鼢鼠的起源进化的研究却很少,并且存在着甘肃鼢鼠属于[url=http://baike.baidu.com/view/113192.htm][color=#000000]瞎鼠科[/color][/url]和仓鼠二者之争,因此,为了更好的阐明甘肃鼢鼠的起源,还需要做更多、更深入的研究。
刚接触这个检测项目,用的马尔文的仪器,请问大家符合药典规定的粒径和zeta电位检测标准粒子用什么,大家购买的什么品牌的,标准粒径和电位是多少?做mRNA纳米脂质颗粒zeta电位检测大家用什么稀释剂,因为没有测物理常数的仪器,所以用不了较优的产品背景溶液,有什么别的稀释剂可以代替,使检测结果与真实值偏差较小。
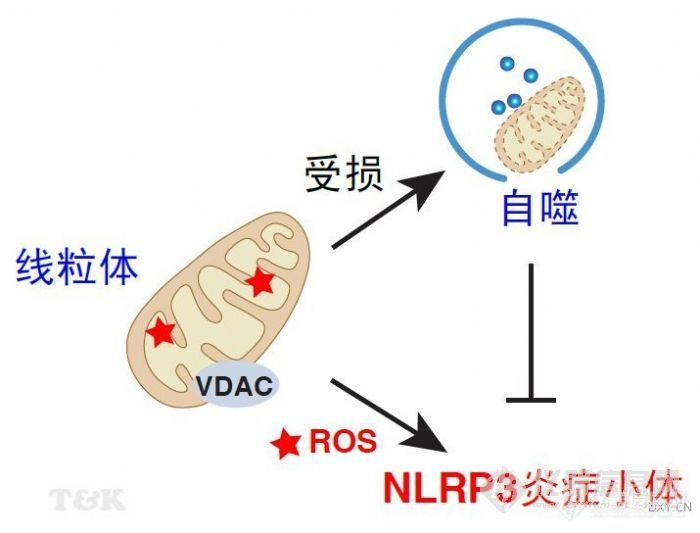
http://ng1.17img.cn/bbsfiles/images/2010/12/201012031408_264152_2019107_3.jpg天然免疫(natural immunity)指个体出生时即具备的免疫能力,是抵抗病原微生物感染的第一道防线。天然免疫主要通过模式识别受体(pattern recognition receptor,PRR)来识别病原体相关分子模式(pathogen-associated molecular pattern,PAMP)。 NLRP3是PRR大家族中Nod样受体亚家族成员之一,在激活之后能够通过与接头蛋白相互作用,活化半胱氨酸蛋白酶-1(caspase-1),形成蛋白复合物“炎症小体(inflammasome)”,进而对Pro-IL-1β等底物进行切割使其成熟并释放到胞外发挥功能。尽管研究发现许多PAMP与DAMP(损伤相关分子模式)能够活化NLRP3炎症小体,但是其调控过程仍不十分清楚。 2010年12月1日,《Nature》杂志发表了来自瑞士科学家Jurg Tschopp研究组的最新成果(参考文献1)。Jurg Tschopp是享誉世界的炎症小体和IL-1研究的科学家,他与Kate Schroder是该领域的领跑者和掌门人,经常见二人合著或分著综述在CNS上(参考文献2,3,11)。这次,Jurg Tschopp研究组发现线粒体来源的活性氧(ROS)是调控NLRP3炎症小体活化的关键信号。同时发现自噬(autophagy)及其线粒体自噬(mitophagy)调控了线粒体的质量,减少受损的线粒体数目,从而防止了ROS诱导的NLRP3炎症小体活化。自噬缺失的细胞加剧了NLRP3炎症小体活化。进一步表明线粒体的电压依赖性阴离子通道蛋白(voltage-dependent anion channel,VDAC)调控了线粒体ROS的产生以及NLRP3炎症小体的活化。本研究对于解释为什么炎症疾病常伴有线粒体损伤提供了新证据。实际上Jurg Tschopp研究组在今年早期的《Nature Immunol》杂志上已发文提示线粒体TXNIP蛋白调控了ROS诱导的NLRP3炎症小体活化(参考文献4)。这次《Nature》杂志的这篇文章进一步将细胞自噬这个国际研究热点和基本的细胞应激机制引入到线粒体介导的NLRP3炎症小体活化。然而,我们应该清楚的看到最早关于自噬是NLRP3炎症小体活化的负调控机制的文章来自日本科学家2008年发表在《Nature》的报道,该报道表明体内外缺失自噬调节基因Atg16L1导致了NLRP3炎症小体活化(参考文献5)。2008年同期《Nature》还报道了Atg16L1在一种胃肠道的慢性非特异性炎症性疾病克罗恩病(Crohn病)的重要作用(参考文献6)。短短两年内,Atg16L1与炎症相关性疾病的基础和临床研究呈爆发式增长。
中科院生物学家通过对比分别基于线粒体基因组和核基因外显子估算的分歧时间,发现基于线粒体基因的分子钟分析严重高估了分歧事件发生的时间。http://ng1.17img.cn/bbsfiles/images/2011/09/201109162159_317239_1609805_3.jpg
可杀死处于所有阶段的疟原虫 新华社华盛顿3月21日电(记者任海军)美国研究人员日前报告说,他们开发出一种新型抗疟疾药物,在动物实验中,其效果强于临床抗疟药,且不易产生耐药性。 这种新药名为ELQ-300。在动物实验中,它能以疟原虫的线粒体为标靶发挥作用,杀死处于所有阶段的寄生虫。疟原虫体内线粒体的主要功能是制造DNA所需的构建模块,而新药能阻断这一过程。 研究人员还发现,新药治疗实验鼠的效果强于临床抗疟药阿托伐醌。仅需阿托伐醌剂量的十分之一,新药便可保护实验鼠免受蚊虫传播的疟原虫感染。此外,研究发现很难筛选出对新药产生耐药性的疟原虫,这意味着新药如果用于人类,可以在临床上应用较长时间。 相关研究报告发表在美国最新一期《科学转化医学》杂志上。领导研究的俄勒冈卫生科学大学教授迈克尔·里斯科表示,这一药物“具有阻止疟疾传播的潜力”,但在用于人类临床试验前还需接受严格的安全测试。 疟疾是由疟原虫引起的疾病,如不治疗,疟疾可能中断对重要器官的供血,从而威胁生命。在世界很多地区,疟原虫已经对一些抗疟药产生耐药性。据统计,全球每年约有近百万人死于疟疾,其中大部分分布在撒哈拉以南非洲地区。
[size=3]据《每日科学》8月17日报道, 美国莱斯大学研究发现:“夏娃”生活于20万年以前。 不要惊讶,该“夏娃”只不过是国外一些科学家提出的一个现代人类起源理论,被称为“夏娃”理论。上世纪80年代,美国加州大学伯克莱分校的威尔逊遗传小组发现:全人类的线粒体DNA基本相同,差异很少,而线粒体DNA又是严格的母系遗传。因此,他们推测,现代世界各种族居民的线粒体DNA最终都是从一个共同的女性祖先那儿遗传下来的。他们还推测:现代人类的线粒体DNA均来自非洲的一位女性,她是人类各种族的共同祖先。 尽管“夏娃”理论在科学界仍曾争议,但毫无疑问的是:线粒体,这个微小的细胞器,正通过控制机体的能量来控制着生命的新陈代谢。但谁又知道,线粒体,原只不过是寄生于人体的细菌而已。 无论在形态、化学组成、物理性质,还是活动状态、遗传体系等方面,线粒体都长得很像细菌。因此,在科学界,有一种种被广泛接受的学说——内共生学说,学说认为:原先独立生活的细菌在真核生物的共同祖先中繁殖,形成今天的线粒体。在这个学说里,线粒体只不过是生命细胞曾经的奴隶。远古时期的某天,某个原始的真核细胞吞噬了一个细菌,这个被吞噬的细菌,逐步进化,形成了今天的线粒体。而且在进化的过程中,这个细菌逐步丧失了独立性,经过许多代的奴役之后,最终变得完全寄人篱下。但在这一过程中,它也将大量遗传信息转移到了细胞里。 [/size]
原创大赛进行得好快啊·~转眼就到9月份了,正好将暑期的实验总结下,来参加大赛了~~~~大家轻拍板砖~~~ 奖品许愿:容我再想几天哈~~~摘 要:本实验用酚—氯仿抽提的经典方法从动物心脏中提取总DNA,电泳检测后进行分离纯化,获得总DNA样品。以D-loop序列为引物,用提取的总DNA作为模板,在适宜条件下PCR快速扩增mtDNA D-loop区,获得了大量的目的片段。 线粒体(mitoch ondria)是真核细胞核外唯一的遗传物质,哺乳动物mtDNA是约16.5kb的闭环双链分子,线粒体DNA的结构上有一个独特的D—Loop环(displacement loop region):位于tRNA—Pro和tRNA—Pile的基因之间,由少数碱基构成一个突出结构。在线粒体DNA上,D—Loop环是整个线粒体基因组序列和长度变异最大的区域,其进化速度最快,一般用于种内种群间的系统进化分析。线粒体作为真核生物胞质遗传的重要组成部分,mtDNA是由卵细胞传递给后代,被认为属于典型的母性遗传。 有关生物进化的研究一直是科学家关注的问题,现在国内外很多学者利用mtDNA D-loop区研究生物进化,采用Neighbor-joining算法,绘制生物系统进化树。哺乳动物线粒体DNA的D-loop区是其复制和转录的起始区域,是一个高度多态性和突变性的区域,其中D—loop重链RNA(DH-RNA)与复制和转录功能密切相关。目前有关mtDNA D-Loop的遗传变异分析的研究已在人、牛、猪、马、鸭、鱼类、昆虫类、两栖类等多种动物中开展。为了探讨动物的分类地位,我们以位于D - loop 两端的tRNApro和12S rRNA 基因内的部分序列设计的2 对引物通过聚合酶链式反应(Polymerase Chain Reaction , PCR) 扩增mtDNA D - loop ( P1 和P2) ,通过测定产物序列,相关比较,进一步分析其分类地位,以及mtDNA D环多态性,为以后进一步研究线粒体D—Loop区的多态性与疾病的关系以及分析物种之间的进化规律奠定了基础。 1 材料与方法 1.1.1实验试剂 提取DNA:生理盐水(去除组织中的淤血)、 TES液(抑制DNA酶作用)、SDS(表面活性剂,抑制DNA酶)、饱和酚(将蛋白质变性)、酚∶氯仿∶异戊醇(体积比为25∶24∶1)、氯仿∶异戊醇(除去酚)、无水乙醇(预冷,沉淀DNA)、预冷的乙醇(洗涤DNA)、 TE(降解DNA)、蛋白酶K(降解蛋白质) 电泳: TBE缓冲液(pH8.0)、TBE缓冲液、琼脂糖(凝胶介质)、溴化乙啶(EB,荧光染料)、溴酚蓝(指示剂)、标准λDNA(48kb,Mark)、上样液(蔗糖+溴酚蓝) PCR扩增:引物P1:5’-TATGTACCATGAGGACAAATATC-3’,P2:5’-ATTACACCTCCTAATTTATTAGGAATC-3’ 、dNTP(2.5mmol·L-1)、10×Buffer溶液、TaqDNA聚合酶(2.5U·μl-1)、超纯水、Marker(DL2000)、PCR产物回收试剂盒等。 1.1.2仪器 微量移液器、Eppendorf离心机、微波炉、DYCp31型电泳仪、凝胶成像系统、恒温水浴锅、PCR仪 1.2 实验方法 1.2.1 总DNA的提取(总的说来采取的步骤图示为:)http://ng1.17img.cn/bbsfiles/images/2010/09/201009021539_241030_1915361_3.jpg 1.2.1.1 将材料解块,用NaCl洗去血污。剪一小块组织,剪碎,放入-20℃预冷的研钵中加入TES充分研磨。 1.2.1.2吸取匀浆液到离心管中,再加入SDS及蛋白酶K(20mg/ml)充分混匀后于56℃保温,每2小时摇一次。放置室温,加入等体积的饱和酚,颠倒混匀呈乳浊液,10000rpm离心10min(液体分层,上层为黄色,下层无色)。分离水相和有机相小心吸取上层含核酸的水相到一个新的离心管中。 1.2.1.3加入等体积酚∶氯仿∶异戊醇(25∶24∶1)380μl,颠倒混匀15min,10000rpm离心10min,取上清液移到一个新的离心管中。再加入等体积氯仿∶异戊醇(24∶1)颠倒混匀15 min。这时液体分层,10000rpm离心10min,取上清液移到一个新的离心管中。 1.2.1.4加入2.5倍体积-20℃的预冷无水乙醇沉淀DNA。于12000rpm离心15min,用乙醇快速洗涤沉淀,离心,弃上清,干燥。加入TE于40C冰箱中保存备用。 1.2.2 电泳检测总DNA 1.2.2.1称取0.24g琼脂糖于三角瓶中加入30ml0.5×TBE缓冲液配成0.8%的胶;用微波炉加热,煮沸,振荡,反复加热振摇2~3次,使糖充分融化。用医用胶布将胶床两端粘好,注意不要超过底部以免放置不平。 1.2.2.2等胶冷却至约60℃时,将融化的琼脂糖小心地倒入已插好梳子的胶床中,再使其自然冷却,直至完全凝固。小心向上方拔出梳子,去掉制胶槽两边的胶带,小心地将胶和胶床放入电泳槽。向电泳槽中加入TBE缓冲液,液面高于胶面1~2mm,以使其全部处于电场内。 1.2.2.3取8μlλDNA,加2μl上样液,混匀后,小心点入上样孔中。取4μl样品DNA,加2μl上样液在parafilm上混匀,同样点入上样孔中。将电压调至100V,接通电源,打开开关,开始电泳。大约20分钟后,待指示剂迁移到距上样孔1.5cm以外时终止电泳,切断电源,取出凝胶,放入含EB的染色液中染色20分钟。凝胶取出后,于凝胶成像系统中观察分析。 1.2.3总DNA的纯化 提取的总DNA加TE补到50μl,加0.5μl的RnaseA,37℃温浴1h;加50μl氯仿:异戊醇,颠倒均匀,大约10次;10000rPm,离心10分钟,取上清夜;加150μl无水乙醇,轻微晃动;12000rPm离心10分钟,弃乙醇;加300μl70%乙醇洗涤DNA,弃乙醇控干,55℃干燥DNA;加25μlTE溶解DNA 1.2.4 PCR扩增线粒体DNAD-LOOP基因 1.2.4.1 引物的设计 扩增引物为昆虫通用引物,引物设计参考Simon等,引物由上海生工生物工程有限公司合成。 1.2.4.2 扩增体系的建立 每一样品的扩增体积均为25μl。将下列试剂按表1的顺序加入0.2mL的EP管中。http://ng1.17img.cn/bbsfiles/images/2010/09/201009021448_241016_1915361_3.jpg 1.2.4.3 PCR扩增 对上述加好试剂的0.2mL的Ep管短暂离心,灭气泡后放入PCR仪。扩增共运行35个循环,每一循环包括:http://ng1.17img.cn/bbsfiles/images/2010/09/201009021550_241036_1915361_3.jpg 扩增产物用1.0%的琼脂糖凝胶电泳检测其大小、纯度及浓度。 1.2.4 PCR扩增产物的电泳检测 称取0.3g琼脂糖于三角瓶中加入30ml的1×TBE缓冲液配成1.0%的凝胶。 从PCR仪中取出已经扩增的样品.用微量取液器取3μl溴酚蓝加入EP管,反复抽吸以混匀,再从中吸取3μl,加入上样孔.吸取6ulMarker点样.插好电极,接通电泳仪电源,调节电压120V,电泳20min,以指示剂迁移位置判断.将凝胶取出置于EB液中染色20min. 凝胶取出后在凝胶成像系统中拍照观察,从中检测其大小、纯度和浓度。 1.2.5 PCR产物的回收、纯化与测序 1.2.5.1 PCR产物的回收 称取0.3g琼脂糖于三角瓶中加入30ml的1×TBE缓冲液配成1.0%的胶。用微量取液器取3μl溴酚蓝加入Ep管,反复抽吸以混匀,再从中吸取25μl,加入上样孔。另外,吸取6μlMark点样。然后插好电极,接通电泳仪电源,调节电压90V或更低,电泳时间1h,将凝胶取出,在紫外灯下观察结果。 在紫外灯下割下目的胶条,按比例加入S1液300μl,50℃水浴10min,使胶块融化,每隔2 min摇匀一次。加100μl的异丙醇,混匀,50℃水浴5 min,混匀之后短暂离心。将溶好的Agarose胶移入吸附柱,12000rpm离心30S,倒掉收集管中的液体。将吸附柱放入同一管中,然后加入W1500μl,12000rpm离心15 S,倒去管中液体,再加入W1500μl后静置1min,离心15S,倒去液体离心1min。 1.2.5.2 纯化 将吸附柱放入一个干净的1.5mlEp管中,加20μl无菌水,37℃保温5 min溶解。 再于12000rpm离心,DNA溶于液体中。 1.2.5.3 PCR回收产物的检测 使用1.0%的胶,用微量取液器取2μl溴酚蓝加入2μlDNA溶液,加入上样孔。另外,吸取4μlMark点样。然后插好电极,接通电泳仪电源,调节电压

http://ng1.17img.cn/bbsfiles/images/2017/01/201701191656_647598_2507958_3.gif药物研发和实时荧光检测技术的应用时间:2014年9月25日 14:30讲师:董文忠先生,Molecular Devices产品经理 毕业于军事医学科学院毒物药物研究所,曾先后就职于安泰医药生物、新医药北京市技术转移中心、药明康德等药物研发机构。长期从事不同靶点、不同技术平台的药物研发和高通量筛选,拥有丰富的药物开发和筛选经验。现担任Molecular Devices产品经理,负责实时高通量荧光检测分析系统和全自动膜片钳系统的应用技术工作。讲座内容: 药物研发是一个漫长且投资巨大的复杂过程,G蛋白偶联受体和离子通道等一直以来作为最主要的药物作用靶点而倍受青睐,所占市场份额超过一半以上。另外,药物的安全性评测的重要性日渐提升并且开始的阶段也越来越早。如何更好、更快、更有效地完成相应的工作?实时高通量荧光检测技术将是您最佳的选择。钙流、膜电位、pH、hERG通道、氯离子、早期心肌毒性检测、cAMP检测等多种应用定能满足您的需求。尊敬的客户,耽误您宝贵的3分钟时间来填写以下webinar注册信息,对于完成注册并当天全程听会的客户,我们会抽取8位听众送出精美小礼品。感谢大家的关注!-------------------------------------------------------------------------------1、报名条件:只要您是仪器网注册用户均可报名参加。2、报名并参会用户有机会获得100元手机充值卡一张哦~3、报名截止时间:2014年9月25日 14:004、报名参会:http://ng1.17img.cn/bbsfiles/images/2014/08/201408011630_508801_2507958_3.jpghttp://ng1.17img.cn/bbsfiles/images/2017/01/201701191656_647598_2507958_3.gif

高效液相色谱法分析苯并芘大鼠肝脏线粒体的代谢产物 本实验建立了一种用高效液相色谱法分析苯并芘及其在大鼠肝脏线粒体中的六种代谢产物的分析方法。使用乙腈、水梯度洗脱作为流动相,紫外探测器分析得到苯并芘的羟基化代谢产物以及苯并芘酮,包括3-羟基苯并芘、9-羟基苯并芘、苯并芘4,5-二氢二醇、苯并芘-7,8-二氢二醇、9,10-二羟基-9,10-二氢苯并芘、苯并芘二酮。其中苯并芘二酮含量最低。该实验结果对于推断细胞CYP1A1酶在体内体外模型中对于苯并芘增毒和解毒作用奠定了重要的基础。 前言:苯并芘是苯与芘稠合而成的一类多环芳烃,苯并芘和其他多环芳烃主要是有机物的不完全燃烧或热解生成,并且在环境中普遍存在。除了污染空气的吸入,摄入的主要途径有吸烟和饮食以及一些职业的摄入如煤、焦炭、沥青的燃烧以及煤焦油的使用。苯并芘能够导致细胞毒性、致畸致突变的毒性以及致癌的毒性。动物实验长期暴露于苯并芘中可导致动物的皮肤、胃、肺组织的癌变。苯并芘在作用于DNA之前需要代谢活化,这也是苯并芘发挥毒性很重要的代谢步骤。细胞色素P450(CYP)酶和环氧化物酶是主要的苯并芘的活化酶,首先CYP酶将苯并芘氧化为环氧化物然后在环氧化物水解酶的作用下生成二氢二醇,CYP同工酶将其进一步的活化为活性成分苯并芘-7,8 - 二氢二醇-9,10 - 环氧化物(BPDE),其可作用于DNA,其优先在鸟嘌呤残基上形成加合物,该加合物是BPDE在体内体外试验中于DNA主要的加合物。在CYP酶中,CYP1A1和B1认为是BaP代谢活化中重要的酶,但是CYP1A1在体内排毒的作用较大于其活化BaP的作用。为了解释这些发现,BaP的体内体外代谢与解毒作用应该进一步进行评价,定性和定量分析BaP在CYP同工酶和环氧化物酶下的所有代谢产物,以及这些致癌物与DNA加成物的评价也很有必要。本实验优选色谱条件使得BaP在大鼠肝脏线粒体内的代谢产物能够很好的分离以及通过紫外检测器灵敏的检测。苯并芘在生物体内的代谢步骤:http://ng1.17img.cn/bbsfiles/images/2014/09/201409291248_516273_2360169_3.jpg材料和方法试剂甲醇(色谱级)乙腈(色谱级),苯并芘 ,NADP+,葡萄糖-6-磷酸,二喹啉甲酸,葡萄糖-6-磷酸脱氢酶微粒体的制备微粒体来自于10只SD大鼠的肝脏,预先用苏丹I处理。微粒体蛋白质浓度通过二辛可宁酸蛋白质测定法测定,牛血清蛋白作对照。CYP同工酶的含量通过示差光谱测定。孵化体系:用于研究BaP代谢的孵化体系包含有100mM磷酸钠缓冲液(pH7.4),NADPH生成体系(1毫NADP+,10mL D-葡萄糖-6 - 磷酸,1U/mL的D-葡萄糖-6-磷酸脱氢酶),0.5mg的微粒体蛋白质,50μM的BaP(溶于5μl甲醇),总体积为500微升。通过加入50μl的NADPH生成体系来启动反应的发生。孵育体系通过未加入酶体系或无NADPH生成体系或无的BaP来控制。孵化在敞开的试管中进行(37℃),20分钟后,取5μl 1mM的非那西丁乙醇溶液加入作为内标物。BaP的代谢物用乙酸乙酯(2×1毫升)萃取两次,并蒸发至干。将样品溶解在25μl的甲醇,通过HPLC分离。BaP代谢物的HPLC分析:安捷伦液相1200高效液相色谱仪配紫外可见检测器,色谱柱为diamonsil 4.6﹡150﹡5u色谱条件:所用的色谱条件如下表: 时间流动相A(乙腈)流动相B(水)流速00%100%0.6ml/min3530%70%4060%40%4580%20%50100%0%我们还对代谢产物进行了质谱
西达本胺通过信号通路调节促进癌细胞凋亡在我国,西达本胺已获批作为PTCL临床用药。西达本胺属于苯酰胺类化合物,是我国自主研发的首个亚型选择性口服HDACI,国家食品药品监督管理局已批准其用于临床试验,其选择性抑制I类HDAC1、2、3亚型和II类HDAC10亚型,可抑制肿瘤细胞增殖、促进凋亡,阻滞周期、引发DNA损伤,还可以增强抗肿瘤免疫反应。与其他抗肿瘤药物相比,西达本胺疗效好、选择性高、不良反应少。西达本胺可激活死亡受体途径和线粒体凋亡途径诱导细胞凋亡,其中最为主要的是线粒体凋亡途径,该途径受Bcl-2家族介导的细胞色素C释放通路调控。抗凋亡蛋白Bcl-2表达受到抑制,促凋亡蛋白Bax表达上调,使线粒体膜电位降低,细胞色素C释放到细胞质中,Caspase途径被激活,细胞发生凋亡。例如:西达本胺增强B淋巴瘤细胞组蛋白H3、H4 乙酰化水平,使线粒体膜电位降低随后激活Caspase 3,促进细胞凋亡;在肾癌中,它可以下调Bcl-2表达,上调Bax表达,随着药物浓度增加引起786-O 细胞凋亡。西达本胺可以调控ROS水平。HDACI可以上调ROS水平,导致DNA双链损伤。研究证明,西达本胺作用于白血病细胞后,诱导细胞内ROS产生,细胞凋亡增加[17]。此外,在胰腺癌细胞系中,西达本胺明显增强细胞内ROS的产生,上调γH2AX(DNA双链断裂的标志物)表达水平,诱发细胞DNA损伤。西达本胺通过调控细胞周期蛋白(Cyclin)、细胞周期蛋白依赖性激酶(Cyclin-dependent kinases,CDKs)以及细胞周期蛋白依赖性激酶抑制剂(Cyclin-dependent kinases inhibition,CDKI)的表达阻滞细胞周期。例如,西达本胺使MM细胞系P21、P27的表达量增高,CDK4、CDK6、Cyclin D2表达量下降,阻滞MM细胞系于G1期[19]。在NK/T细胞淋巴瘤中,西达本胺上调P21表达,下调Cyclin E表达,诱导细胞发生G0/G1期阻滞,从而抑制细胞的增殖。
[size=15px][font=宋体]茯苓([/font][i][font=&]Poria cocos[/font][/i][font=宋体],[/font][font=&]PC[/font][font=宋体])是一种药食同源的真菌,在中国作为中药已有两千多年的使用历史,其化学成分主要包括三萜、多糖、蛋白质和氨基酸,具有抗炎、免疫调节、抗癌、抗高血糖和调节血脂等作用。[/font][font=&][/font][/size][size=15px][font=宋体]代谢功能障碍相关脂肪肝([/font][font=&]MAFLD[/font][font=宋体])是最常见的慢性肝病,目前尚无缓解[/font][font=&]MAFLD[/font][font=宋体]的特定治疗方法。已有研究报道[/font][font=&]PC[/font][font=宋体]还有潜在的抗[/font][font=&]MAFLD[/font][font=宋体]作用,但其抗[/font][font=&]MAFLD[/font][font=宋体]的生物活性成分仍然未知,且[/font][font=&]PC[/font][font=宋体]是否通过调节线粒体功能来缓解[/font][font=&]MAFLD[/font][font=宋体]也不清楚。[/font][/size][size=15px][font=宋体]利用[/font][font=&]L02[/font][font=宋体]肝细胞模型和高脂饮食诱导的[/font][font=&]MAFLD[/font][font=宋体]大鼠模型,发现[/font][font=&]PC [/font][font=宋体]对抗[/font][font=&] MAFLD [/font][font=宋体]的功效。机制上,[/font][font=&]PC[/font][font=宋体]维持线粒体的超微结构,减轻线粒体的氧化应激,调节能量代谢和脂肪酸[/font][font=&]β[/font][font=宋体]氧化,从而减轻脂肪乳剂诱导的细胞脂肪变性和[/font][font=&]HFD[/font][font=宋体]诱导的[/font][font=&]MAFLD[/font][font=宋体]。此外,研究从[/font][font=&]PC[/font][font=宋体]中鉴定出的[/font][font=&]15[/font][font=宋体]种生物活性物质可能是调节线粒体功能以减轻[/font][font=&]MAFLD[/font][font=宋体]的主要有效[/font][font=&]PC[/font][font=宋体]成分,因此,[/font][font=&]PC[/font][font=宋体]可能是预防[/font][font=&]MAFLD[/font][font=宋体]的有前途的线粒体调节剂。[/font][/size] [img=,690,395]https://ng1.17img.cn/bbsfiles/images/2024/09/202409101326367319_3570_6561489_3.png!w690x395.jpg[/img] [size=15px][b][font=&][color=#0070c0]1[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]PC[/color][/font][font=宋体][color=#0070c0]提取物对脂肪乳诱导的脂肪细胞的影响[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]作者首先通过体外细胞实验发现,[/font][font=&]PC[/font][font=宋体]提取物抑制了脂肪乳剂诱导的[/font][font=&]L02[/font][font=宋体]细胞脂肪变性[/font][font=宋体],且[/font][font=&]PC[/font][font=宋体]提取物阻止了线粒体结构和功能的损伤并增强了线粒体的自噬,从而减轻了[/font][font=&]L02[/font][font=宋体]细胞脂肪变性[/font][font=宋体]此外,[/font][font=&]PC [/font][font=宋体]提取物促进脂肪酸[/font][font=&]β [/font][font=宋体]氧化,从而减轻脂肪乳剂诱导的[/font][font=&] L02 [/font][font=宋体]细胞脂肪变性[/font][/size][align=center][size=15px][b][font=&][color=#0070c0]2[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]PC[/color][/font][font=宋体][color=#0070c0]提取物对[/color][/font][font=&][color=#0070c0]HFD[/color][/font][font=宋体][color=#0070c0]喂养大鼠的影响[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]作者接着通过体内实验研究了[/font][font=&]PC[/font][font=宋体]提取物的功能,发现[/font][font=&]HFD[/font][font=宋体]喂养[/font][font=&]12[/font][font=宋体]周后,肝脏指数显著增加,而[/font][font=&]PC[/font][font=宋体]提取物减轻了体重增加和肝肿大。此外,[/font][font=&]PC[/font][font=宋体]提取物可以缓解[/font][font=&]HFD[/font][font=宋体]引起的血脂异常[/font][font=宋体],减轻[/font][font=&]HFD[/font][font=宋体]喂养大鼠的的肝脏脂质异常和肝脏损伤[/font][font=宋体]同样,体内实验表明[/font][font=&]PC[/font][font=宋体]提取物促进线粒体功能,减轻[/font][font=&]MAFLD[/font][font=宋体],并通过刺激脂肪酸[/font][font=&] β [/font][font=宋体]氧化以减轻[/font][font=&]HFD[/font][font=宋体]喂养大鼠的肝脏脂肪变性[/font][/size][/align][align=center] [/align][size=15px][b][font=&][color=#0070c0]3[/color][/font][font=宋体][color=#0070c0]、肝线粒体[/color][/font][font=&][color=#0070c0]PC[/color][/font][font=宋体][color=#0070c0]衍生成分对脂肪细胞的影响[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]接着作者获得[/font][font=&]PCME [/font][font=宋体](茯苓提取物高剂量组的肝线粒体提取物),发现[/font][font=&]PCME[/font][font=宋体]处理后,细胞内脂质积累受到显著抑制,相反,[/font][font=&]MME[/font][font=宋体](模型组肝线粒体提取物)处理未能抑制脂质积累。结果表明来自肝线粒体提取物的[/font][font=&]PC[/font][font=宋体]衍生成分可修复氧化应激和能量代谢,从而减轻细胞脂肪变性[/font][font=宋体]。[/font][/size][font=宋体][size=15px]通过[/size][/font][font=&][size=15px]UHPLC/MS[/size][/font][font=宋体][size=15px]从经[/size][/font][font=&][size=15px]PC[/size][/font][font=宋体][size=15px]处理的大鼠肝线粒体提取物中鉴定出[/size][/font][font=&][size=15px]15[/size][/font][font=宋体][size=15px]种[/size][/font][font=&][size=15px]PC[/size][/font][font=宋体][size=15px]衍生化合物,包括[/size][/font][font=&][size=15px]14[/size][/font][font=宋体][size=15px]种原型化合物([/size][/font][font=&][size=15px]HTA, PAG, DMA,DTA, DPA, DEA, PPAC[/size][/font][font=宋体][size=15px]等)和[/size][/font][font=&][size=15px]1[/size][/font][font=宋体][size=15px]种代谢物(其原型为茯苓酸,[/size][/font][font=&][size=15px]Hydroxypachymicacid[/size][/font][font=宋体][size=15px]),其中有[/size][/font][font=&][size=15px]14[/size][/font][font=宋体][size=15px]种三萜酸和[/size][/font][font=&][size=15px]1[/size][/font][font=宋体][size=15px]种甾醇[/size][/font][font=宋体][size=15px]。[/size][/font][align=center] [/align][size=15px][b][font=&][color=#0070c0]4[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]PC[/color][/font][font=宋体][color=#0070c0]衍生成分和线粒体成分组对脂肪细胞的影响[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]经[/font][font=&]7[/font][font=宋体]种[/font][font=&]PC[/font][font=宋体]衍生单体([/font][font=&]HTA[/font][font=宋体]、[/font][font=&]PAG[/font][font=宋体]、[/font][font=&]DMA[/font][font=宋体]、[/font][font=&]DTA[/font][font=宋体]、[/font][font=&]DPA[/font][font=宋体]、[/font][font=&]DEA[/font][font=宋体]和[/font][font=&]PPAC[/font][font=宋体])分别处理后[/font][font=&], TC[/font][font=宋体]和[/font][font=&]TG[/font][font=宋体]含量显著降低[/font][font=&], SOD[/font][font=宋体]、[/font][font=&]GSH[/font][font=宋体]、[/font][font=&]Na + -K + -ATPase[/font][font=宋体]、[/font][font=&]Ca 2+ -Mg 2+-ATPase[/font][font=宋体]和[/font][font=&]Complex-[/font][font=宋体]Ⅱ含量显著升高[/font][font=&], Complex-I[/font][font=宋体]含量呈上升趋势。这些结果表明[/font][font=&]PC[/font][font=宋体]衍生成分调节脂肪变性肝细胞的氧化应激和能量代谢,从而减少脂质蓄积(图[/font][font=&]9[/font][font=宋体])。[/font][font=&]7[/font][font=宋体]种[/font][font=&]PC[/font][font=宋体]衍生单体组成的[/font][font=&]PCMM ([/font][font=宋体]线粒体[/font][font=&]PC[/font][font=宋体]衍生单体群)处理后细胞[/font][font=&]TC[/font][font=宋体]和[/font][font=&]TG[/font][font=宋体]含量显著降低,[/font][font=&]SOD[/font][font=宋体]、[/font][font=&]GSH[/font][font=宋体]、[/font][font=&]Na+ -K+-ATPase[/font][font=宋体]、[/font][font=&]Ca2+ -Mg2+ -ATPase[/font][font=宋体]、[/font][font=&]Complex-[/font][font=宋体]Ⅱ含量显著升高,[/font][font=&]Complex-[/font][font=宋体]Ⅰ含量呈上升趋势,逆转脂肪乳刺激后细胞[/font][font=&]ACADL[/font][font=宋体]、[/font][font=&]ECHS[/font][font=宋体]、[/font][font=&]PPAR-α[/font][font=宋体]的[/font][font=&]mRNA[/font][font=宋体]表达水平的降低。这些结果表明[/font][font=&]PCMM[/font][font=宋体]可以改善氧化应激和能量代谢,并调控脂肪酸[/font][font=&]β[/font][font=宋体]氧化相关基因的表达,从而减轻细胞脂肪变性[/font][/size]
[size=15px][font=宋体]帕金森病([/font][font=&]PD[/font][font=宋体])是一种神经退行性疾病,线粒体功能障碍参与帕金森病的发病机制。心磷脂([/font][font=&]CL[/font][font=宋体])是线粒体动力学和功能中的关键磷脂,在塑造和维持蛋白质[/font][font=&]-[/font][font=宋体]蛋白质和蛋白质[/font][font=&]-[/font][font=宋体]膜相互作用方面起着关键作用,同时还起着稳定线粒体呼吸链的作用。许多神经系统疾病都与[/font][font=&]CL[/font][font=宋体]异常有关。鉴于[/font][font=&] CL[/font][font=宋体]、线粒体和神经系统疾病之间存在相关性,保持线粒体[/font][font=&]CL[/font][font=宋体]含量并维持其正常分布的小分子在预防和减轻线粒体损伤方面具有前景。[/font][font=&][/font][/size][size=15px][font=宋体]人参皂苷[/font][font=&]Rg3[/font][font=宋体]([/font][font=&]Rg3[/font][font=宋体])是一种源自人参的突出生物活性化合物,在多种恶性肿瘤中表现出显著的抗肿瘤功效,并发挥以其抗氧化和抗炎特性为特征的神经保护作用。在临床前模型中,[/font][font=&]Rg3[/font][font=宋体]在鱼藤酮诱导的[/font][font=&]PD[/font][font=宋体]小鼠中表现出运动功能的增强。[/font][font=&]Rg3[/font][font=宋体]的神经保护机制部分涉及其抗氧化特性,以及线粒体通透性过渡孔张口的抑制。然而,[/font][font=&]Rg3[/font][font=宋体]介导其作用的特定蛋白质靶点仍有待阐明。[/font][/size][size=15px][font=宋体]人参皂苷[/font][font=&]Rg3[/font][font=宋体]([/font][font=&]Rg3[/font][font=宋体])能够提高心磷脂水平,进而通过增强线粒体未折叠蛋白反应、促进线粒体自噬和增强线粒体氧化磷酸化来促进线粒体稳态,增强酪氨酸羟化酶阳性([/font][font=&]TH[/font][font=宋体])多巴胺能神经元的存活率,从而改善了[/font][font=&]PD[/font][font=宋体]小鼠模型中的运动性能。机制上,[/font][font=&]GRB2[/font][font=宋体]蛋白是[/font][font=&] Rg3 [/font][font=宋体]的直接分子靶标,[/font][font=&]Rg3[/font][font=宋体]促进[/font][font=&]GRB2[/font][font=宋体]和[/font][font=&]TRKA[/font][font=宋体]的互作,增加[/font][font=&]CRLS1[/font][font=宋体](心磷脂合酶[/font][font=&] 1[/font][font=宋体])基因表达并促进心磷脂合成。此外,研究还发现富马酸替诺福韦二吡呋酯([/font][font=&]TDF[/font][font=宋体])同样可以诱导的[/font][font=&]GRB2[/font][font=宋体]和[/font][font=&]TRKA[/font][font=宋体]之间的增强结合促进[/font][font=&]CL[/font][font=宋体]的合成和恢复线粒体稳态,显示出预防[/font][font=&]PD[/font][font=宋体]的潜力。 [size=15px][/size][size=15px][b]1、PD模型中的CL耗竭和线粒体功能障碍[/b][/size][size=15px]作者首先通过体外和体内PD模型中评估了CL水平,发现用A53T-αSyn或MPTP处理的小鼠的SN表现出线粒体CL水平显著降低,转染pCMV3-A53T-αSyn-His的小鼠线粒体CL水平显著较低。此外,用mpp[i][/i]或6-OHDA处理的人神经母细胞瘤细胞系SH-SY5Y的线粒体CL水平较低,这些发现表明PD患者的CL水平降低。接着评估PD中的线粒体功能,发现线粒体CL水平降低及导致的线粒体功能障碍在PD的发病机制中起着关键作用[/size] [size=15px][/size][size=15px][b]2、[/b][/size][size=15px][b]Rg3[/b][/size][size=15px][b]通过GRB2上调CL以提供神经保护[/b][/size][size=15px]接着,作者制定了一种筛选策略,旨在识别能够恢复CL水平的化合物以预防PD。通过10-壬基吖啶橙(NAO)荧光筛选包含包含400多种天然产物的化合物库,发现连翘苷、人参皂苷Rg3、紫云英苷效果最佳。利用秀丽隐杆线虫评估这三种化合物在清除αSyn聚集体中的功效,发现Rg3表现出最显著的效果。此外,Rg3给药促进mitoUPR水平恢复,表明Rg3是一种有效的抗PD化合物,可提高CL水平并维持线粒体稳态。Rg3的功效在哺乳动物神经细胞中也得到证实,且在PD小鼠模型中,Rg3给药阻止了A53T-αSyn诱导的黑质DA神经元的丢失。上述结果结合一些已有报道表明Rg3 通过增强线粒体 CL 水平和维持线粒体稳态来赋予神经保护作用。然而,Rg3控制CL的确切机制尚不清楚。[/size][size=15px]接着作者采用有限蛋白水解法(LiP-SMap)鉴定Rg3的直接靶点,通过FC值和p值筛选确定13 种候选蛋白,通过siRNA在SH-SY5Y 细胞中敲低13个候选基因,发现Rg3对A53T-αSyn诱导的神经损伤的保护作用仅在GRB2沉默后完全消除。线粒体CL的直接定量表明Rg3处理恢复了A53T-αSyn诱导的线粒体CL水平降低,当GRB2沉默时,这种效应消失。接着作者通过CETSA、BLI技术确定Rg3直接与GRB2结合,分子对接预测了结合模式[/size] [size=15px][/size][size=15px][b]3、Rg3增强人神经细胞中GRB2与TRKA的相互作用[/b][/size][size=15px]鉴于GRB2直接与NTRK1或TRKA结合对EGFR信号传导至关重要,作者研究了Rg3是否以TRKA或EGFR依赖性方式保护SH-SY5Y细胞。结果显示,用 Rg3 处理促进了GRB2和TRKA之间的互作,且线粒体CL的直接定量显示Rg3处理恢复了A53T-αSyn诱导的线粒体CL水平降低,当TRKA沉默时,这种效应消失。此外,分子对接和SPR分析表明Rg3与GRB2的互作导致TRKA和GRB2之间的结合增强。GRB2与TRKA的细胞内结构域结合,从而启动TRKA激活并促进其磷酸化[/size] [size=15px][/size][size=15px][b]4、Rg3诱导人神经细胞中CRLS1表达上调[/b][/size][size=15px]CL通过一系列酶进行合成和代谢。为了确定在维持PD中CL水平方面至关重要的酶,作者采用了RNAi技术来沉默每个候选基因,发现只有敲低CRLS1才能完全消除线虫中的Rg3效应。此外,CRLS1敲低降低CL水平,而在PD模型中,CL补充逆转了这些效应。结果表明,Rg3通过激活CRLS1来提高CL水平,以减轻PD模型中的神经元损失。通过查阅荷兰脑库的数据发现PD患者和对照个体之间的SN中,CRLS1基因的表达显著下降。在细胞模型中同样发现CRLS1 表达显著降低。当CRLS1沉默后,Rg3对A53T-αSyn诱导的神经损伤的保护作用完全消除。此外,Rg3处理增强了 SH-SY5Y 细胞中 CRLS1 的表达,而这些细胞中GRB2或TRKA的敲低减弱了Rg3诱导的表达,表明Rg3通过GRB2和TRKA激活CRLS1表达。进一步研究发现,Rg3激活TRKA-GRB2-ERK-CRLS1通路,协调刺激CL生物合成以维持线粒体稳态[/size] [size=15px][/size][size=15px][b]5、Rg3以EVI1依赖性方式介导CRLS1上调[/b][/size][size=15px]为了阐明 CRLS1 转录上调的机制,作者使用TRANSFAC的匹配工具预测了转录因子(TFs)和相关的TF结合位点。在CRLS1的启动子区域中鉴定出4个推定的 TF,使用RNAi技术SH-SY5Y细胞中敲低这些TFs,发现在A53T-αSyn存在的情况下,只有EVI1敲低消除了Rg3的神经保护作用。深入研究表明表明 Rg3 协调 TRKA-GRB2-ERK 通路的激活,最终导致 EVI1 的核易位和 CRLS1 转录的激活[/size] [size=15px][/size][size=15px][b]6、Rg3通过Grb2和Crls1缓解A53T-αSyn诱导的运动功能缺陷[/b][/size][size=15px]运动功能障碍是人类患者和小鼠模型中PD的主要病理表现。作者利用两种不同的PD小鼠模型深入探讨了Rg3的保护作用,发现A53T-αSyn 或 MPTP 处理的小鼠在Rg3治疗后运动缺陷显著改善。而在小鼠SN中敲低Grb2或Crls1后,Rg3的神经保护功效完全消失。此外,脂质组学显示Rg3 治疗增加了正常和 A53T-αSyn 诱导的 PD 小鼠 SN 中的 CL 水平,而小鼠SN中Grb2或Crls1的敲低逆转了Rg3对CL水平的影响。这些结果表明Grb2和Crls1在Rg3对小鼠黑质DA神经元的保护作用中起着关键作用[/size] [/font][/size][align=center][font=宋体][size=15px][color=black][/color][/size][/font][/align][size=15px][b][font=&][color=#0070c0]7[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]Rg3[/color][/font][font=宋体][color=#0070c0]和[/color][/font][font=&][color=#0070c0]L-DOPA[/color][/font][font=宋体][color=#0070c0]在缓解[/color][/font][font=&][color=#0070c0]A53T-αSyn[/color][/font][font=宋体][color=#0070c0]诱导的运动功能缺陷中的协同作用[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体][color=black]左旋多巴是临床实践中为[/color][/font][font=&][color=black]PD[/color][/font][font=宋体][color=black]患者开出的标准药物,鉴于[/color][/font][font=&][color=black]Rg3[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]L-DOPA[/color][/font][font=宋体][color=black]之间的作用机制不同,作者假设它们的同时给药可能会产生增强对运动功能障碍和神经退行性变的保护作用。结果显示,左旋多巴单药治疗相比,[/color][/font][font=&][color=black]Rg3[/color][/font][font=宋体][color=black]和左旋多巴的联合给药在运动功能和神经保护作用方面表现出显著的改善。这些发现表明[/color][/font][font=&][color=black]Rg3[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]L-DOPA[/color][/font][font=宋体][color=black]的联合使用代表了一种新颖且有前途的[/color][/font][font=&][color=black]PD[/color][/font][font=宋体][color=black]临床预防策略[/color][/font][/size] [size=15px][font=宋体][color=black][/color][/font][font=&][color=black][/color][/font][/size][size=15px][b][font=&][color=#0070c0]8[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=宋体][color=#0070c0]靶向[/color][/font][font=&][color=#0070c0]TRKA-GRB2-CRLS1[/color][/font][font=宋体][color=#0070c0]轴的[/color][/font][font=&][color=#0070c0]TDF[/color][/font][font=宋体][color=#0070c0]增强了[/color][/font][font=&][color=#0070c0]CL[/color][/font][font=宋体][color=#0070c0]表达缓解[/color][/font][font=&][color=#0070c0]PD[/color][/font][font=宋体][color=#0070c0]症状[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体][color=black]为了进一步验证[/color][/font][font=&][color=black]TRKA-GRB2[/color][/font][font=宋体][color=black]轴靶向分子在增强[/color][/font][font=&][color=black]CL[/color][/font][font=宋体][color=black]表达和改善[/color][/font][font=&][color=black]PD[/color][/font][font=宋体][color=black]方面的普遍影响,作者采用了表面等离子体共振([/color][/font][font=&][color=black]SPR[/color][/font][font=宋体][color=black])技术筛选与[/color][/font][font=&][color=black]GRB2[/color][/font][font=宋体][color=black]结合的化合物,鉴定鉴定了[/color][/font][font=&][color=black]11[/color][/font][font=宋体][color=black]种化合物。其中,[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]和丁酸氯倍他松处理后,暴露于[/color][/font][font=&][color=black]mpp+[/color][/font][font=宋体][color=black]或[/color][/font][font=&][color=black]6-OHDA[/color][/font][font=宋体][color=black]的[/color][/font][font=&][color=black]SH-SY5Y[/color][/font][font=宋体][color=black]细胞存活率增加。[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]是替诺福韦的前药,替诺福韦是一种核苷酸逆转录酶抑制剂,丁酸氯倍他松是专门为眼科类固醇治疗配制的,考虑到其口服给药能力,作者选择了[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]进行进一步研究。结果显示,[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]促进[/color][/font][font=&][color=black]SH-SY5Y [/color][/font][font=宋体][color=black]细胞中过表达的[/color][/font][font=&][color=black]A53T-αSyn[/color][/font][font=宋体][color=black]的清除,并促进[/color][/font][font=&][color=black]GRB2[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]TRKA[/color][/font][font=宋体][color=black]之间的互作。此外,[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]处理增强[/color][/font][font=&][color=black]CRLS1[/color][/font][font=宋体][color=black]表达,并恢复了[/color][/font][font=&][color=black]SH-SY5Y[/color][/font][font=宋体][color=black]细胞中由[/color][/font][font=&][color=black]A53T-αSyn[/color][/font][font=宋体][color=black]引起的线粒体[/color][/font][font=&][color=black]CL[/color][/font][font=宋体][color=black]水平降低。然而,在[/color][/font][font=&][color=black]SH-SY5Y[/color][/font][font=宋体][color=black]细胞中敲低[/color][/font][font=&][color=black]GRB2[/color][/font][font=宋体][color=black]或[/color][/font][font=&][color=black]TRKA[/color][/font][font=宋体][color=black]减弱了[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]诱导的效应。此外,[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]增加了[/color][/font][font=&][color=black]TRKA[/color][/font][font=宋体][color=black]磷酸化、[/color][/font][font=&][color=black]ERK[/color][/font][font=宋体][color=black]磷酸化和[/color][/font][font=&][color=black]CRLS1[/color][/font][font=宋体][color=black]的表达。动物实验显示[/color][/font][font=&][color=black]TDF[/color][/font][font=宋体][color=black]治疗后后运动能力的损害,以及[/color][/font][font=&][color=black]A53T-αSyn[/color][/font][font=宋体][color=black]诱导的黑质[/color][/font][font=&][color=black]DA[/color][/font][font=宋体][color=black]神经元的损失得到缓解([/color][/font][/size] [size=15px][font=宋体][color=black][/color][/font][font=&][color=black][/color][/font][/size][size=15px][b][font=宋体][color=#0070c0]总结[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体][color=black]研究通过多个模型的综合筛选和验证,发现人参皂苷[/color][/font][font=&][color=black]Rg3[/color][/font][font=宋体][color=black]([/color][/font][font=&][color=black]Rg3[/color][/font][font=宋体][color=black])能够提高心磷脂水平。心磷脂水平的这种增加通过增强线粒体未折叠蛋白反应、促进线粒体自噬和增强线粒体氧化磷酸化来促进线粒体稳态,改善了[/color][/font][font=&][color=black] PD [/color][/font][font=宋体][color=black]小鼠模型中的运动性能。通过靶点筛选和验证技术发现[/color][/font][font=&][color=black]GRB2[/color][/font][font=宋体][color=black]是[/color][/font][font=&][color=black] Rg3[/color][/font][font=宋体][color=black]的分子靶标,[/color][/font][font=&][color=black]Rg3 [/color][/font][font=宋体][color=black]促进[/col
[size=15px][font=&][font=宋体]对乙酰氨基酚([/font][font=&]APAP[/font][font=宋体])过量是药物性肝损伤的主要原因。[/font][font=&]Sirtuins 5[/font][font=宋体]([/font][font=&]SIRT5[/font][font=宋体])与各种肝脏疾病的发展有关。然而,其在[/font][font=&] APAP [/font][font=宋体]诱发的急性肝损伤([/font][font=&]AILI[/font][font=宋体])中的作用仍不清楚。[/font]SIRT5[/font][font=宋体]在[/font][font=&]AILI[/font][font=宋体]中显著下调,并且[/font][font=&]SIRT5[/font][font=宋体]耗竭加剧了体内和体外的线粒体氧化应激。从机制上讲,[/font][font=&]SIRT5[/font][font=宋体]在对乙醛脱氢酶[/font][font=&]2[/font][font=宋体]([/font][font=&]ALDH2[/font][font=宋体])的[/font][font=&]K385[/font][font=宋体]位点进行去琥珀酰化,从而保持[/font][font=&]ALDH2[/font][font=宋体]的酶活性,进而抑制炎症和线粒体氧化应激。此外,[/font][/size][font=宋体][size=15px]研究发现葛根素([/size][/font][font=&][size=15px]puerarin[/size][/font][font=宋体][size=15px])可促[/size][/font][size=15px][font=宋体]进[/font][font=&]SIRT5[/font][font=宋体]去琥珀酰化酶活性并缓解[/font][font=&]AILI[/font][font=宋体]。 [size=15px][b]1、AILI 中肝细胞SIRT5表达显著下调[/b][/size] [size=15px]作者首先通过RNA测序发现APAP[/size][font=宋体]处理[/font][size=15px]后,肝脏组织中 SIRT5 表达显著下调。进一步验证SIRT5参与AILI,发现APAP处理的小鼠血清ALT和AST水平均不同程度升高,且q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]、Western blott和免疫组化检测显示APAP处理后肝脏中SIRT5下调,表明SIRT5是AILI发展的关键介质 [/size][/font][/size][b][font=&][color=#0070c0]2[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]SIRT5 [/color][/font][font=宋体][color=#0070c0]改善[/color][/font][font=&][color=#0070c0]APAP[/color][/font][font=宋体][color=#0070c0]诱导的肝毒性[/color][/font][/b][size=15px][font=宋体][size=15px] [/size] [size=15px]作者构建了SIRT5-KO小鼠和AAV介导的肝脏特异性SIRT5过表达小鼠,以进一步研究SIRT5在AILI中的作用。结果显示APAP处理后WT小鼠血清ALT和AST水平显著升高,且SIRT5-KO小鼠的血清ALT和AST水平升高更为明显,肝脏坏死显著加重,肝细胞死亡率更高,而SIRT5过表达显著改善APAP引起的肝脏损伤,肝细胞死亡率显著降低,结果表明 SIRT5 可减轻 APAP 诱导的肝毒性 [/size] [size=15px][b]3、SIRT5抑制APAP诱导的肝脏炎症[/b][/size] [size=15px]多项研究表明APAP 引起的肝毒性与炎症密切相关。作者发现接受APAP处理的SIRT5-KO小鼠CD11b和Ly6g阳性炎症细胞数量显著增加,肝脏中炎症细胞因子的水平显著升高,且NF-κB 信号的激活增加,而肝脏特异性SIRT5过表达小鼠则相反,这些结果表明SIRT5可抑制APAP诱导的AILI肝脏炎症 [/size] [size=15px][b]4、SIRT5 抑制AILI 中APAP诱导的线粒体氧化应激[/b][/size] [size=15px]在AILI过程中,细胞色素P450酶产生过量的毒性反应代谢物NAPQI,消耗GSH并与线粒体蛋白共价结合形成APAP加合物,导致线粒体功能障碍、ROS产生和线粒体细胞死亡因子的释放,最终导致肝细胞死亡。作者研究了SIRT5 KO或过表达对APAP诱导的线粒体氧化应激的影响,体内和体外实验结果表明SIRT5抑制了AILI期间的线粒体氧化应激 [/size] [size=15px][b]5、SIRT5缺乏导致AILI中蛋白质琥珀酰化全面增加[/b][/size] [size=15px]鉴于SIRT5在去琥珀酰化中的作用明确,作者采用[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]/MS分析了APAP处理的WT和SIRT5-KO小鼠肝脏中的琥珀酰化。结果显示共有465种蛋白质中的953个位点表现出差异琥珀酰化,其中359种蛋白质中的802个位点显示琥珀酰化水平增加,而106种蛋白质中的151个位点显示琥珀酰化水平降低,结果表明SIRT5缺陷导致AILI中蛋白质琥珀酰化整体增加,这在体内和体外得到了进一步的验证 [/size] [size=15px][b]6、SIRT5在K385残基处使ALDH2去琥珀酰化[/b][/size] [size=15px]SIRT5缺乏导致参与线粒体氧化应激的关键酶ALDH2的琥珀酰化显著上调。进一步探索SIRT5调控ALDH2琥珀酰化的具体分子机制,免疫荧光发现SIRT5与ALDH2共定位,免疫共沉淀实验表明SIRT5与ALDH2互作,且SIRT5敲除显著上调了体内和体外ALDH2的琥珀酰化水平,但对ALDH2的总蛋白浓度没有影响,相反SIRT5过表达显著降低ALDH2的琥珀酰化水平。进一步检测发现SIRT5缺乏会抑制ALDH2的酶活性,而SIRT5过表达会增加ALDH2的活性。[/size] [size=15px][url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]/MS显示ALDH2中三个位点(K370、K377、K385)琥珀酰化显著增加,其中K370和K385在不同物种中高度保守,作者通过将赖氨酸(K)突变为谷氨酸(E)模拟琥珀酰化,将K突变为精氨酸(R)模拟去琥珀酰化,发现K385是ALDH2上的关键琥珀酰化位点,且K385而非K370的琥珀酰化影响ALDH2的酶活性。此外,SIRT5主要通过对ALDH2在K385残基上的去琥珀酰化来减轻AILI [/size] [size=15px][b]7、ALDH2在K385残基处的去琥珀酰化可保护小鼠免受 AILI的侵害[/b][/size] [size=15px]为了研究ALDH2-K385去琥珀酰化在AILI中的作用,作者建立了AAV-GFP、AAV-ALDH2-WT和AAV-ALDH2-385K-E过表达转染小鼠,并对其进行APAP处理。结果显示APAP 给药增加ALDH2的琥珀酰化,而ALDH2-385K-E小鼠肝脏中ALDH2的琥珀酰化程度低于ALDH2-WT小鼠。此外,在APAP给药后,ALDH2-385K-E小鼠的转氨酶水平、肝坏死面积和肝细胞死亡增加,线粒体氧化应激和炎症加重。数据表明ALDH2在K385的去琥珀酰化可保护小鼠免受AILI的侵害 [/size] [size=15px][b]8、[/b][/size][size=15px][b]K385 [/b][/size][size=15px][b]位点ALDH2去琥珀酰化介导SIRT5对AILI的保护作用[/b][/size] [size=15px]为了研究SIRT5对ALDH2去琥珀酰化在体内AILI中的作用,作者通过尾静脉注射表达 AAV-GFP、AAV-ALDH2-WT或AAV-ALDH2-385K-E的相关AAV,在SIRT5-KO小鼠中过表达各种形式的ALDH2,这些小鼠随后接受APAP治疗。结果显示SIRT5缺乏显著升高血清转氨酶水平,在APAP处理后引起坏死和肝细胞死亡,而 ALDH2-WT的过表达显著改善了肝损伤。此外,ALDH2-WT小鼠的肝脏氧化和炎症明显减少,但ALDH2-385KE小鼠的肝脏氧化和炎症没有减少,数据表明ALDH2在K385处的去琥珀酰化介导了SIRT5对AILI的保护作用 [/size][size=15px][b]9、葛根素促进SIRT5减轻AILI[/b][/size] [size=15px]为探究SIRT5激动剂对AILI的治疗作用,作者通过虚拟筛选寻找能与SIRT5结合的天然化合物。根据对接结果筛选出10个亲和能最低的化合物,进一步考察其对SIRT5去琥珀酰化酶活性的影响,其中葛根素对SIRT5去琥珀酰化酶活性的提高最为显著。分子对接分析显示SIRT5能与葛根素结合,分子动力学模拟在原子水平上证实了SIRT5-葛根素复合物的结合稳定性和动力学。接着在体内验证了葛根素对APAP诱导的肝损伤的影响,发现葛根素组在APAP刺激后血清AST和ALT水平降低,肝脏坏死和肝细胞死亡减少,APAP 诱导的氧化应激和炎症明显被抑制。结果表明葛根素通过药理学激活SIRT5减轻AILI,提示葛根素是临床治疗AILI的一种有前途的药物[/size][/font][/size]
[img=,690,690]https://ng1.17img.cn/bbsfiles/images/2024/06/202406200955496234_5255_5604214_3.jpg!w690x690.jpg[/img] 土壤氧化还原电位检测仪是一种专门用于测量土壤氧化还原电位的专业设备,具有众多显著的优点。这些优点不仅体现在其测量技术的先进性,还体现在其功能的多样性、操作的简便性以及数据记录和分析的完善性上。 首先,土壤氧化还原电位检测仪采用了高精度的测量技术,这使得它能够快速、准确地测量土壤中的氧化还原电位。这种高精度的测量技术,可以捕捉到土壤氧化还原电位微小的变化,从而为农业生产、环境保护以及科学实验等领域提供准确的数据支持。 其次,土壤氧化还原电位检测仪具有多功能性。它可以测量多种类型的土壤,无论是湿润土壤还是新鲜土壤,都可以进行精确的测量。同时,该仪器还可以测试土壤的pH值、温度等其他相关参数,为用户提供了全面的土壤信息。这种多功能性使得土壤氧化还原电位检测仪能够适应不同的应用场景,满足不同用户的需求。 再者,土壤氧化还原电位检测仪的操作简便性也是其一大优点。用户只需将传感器插入土壤中并连接到仪器,然后按下启动按钮即可开始测量。这种简单的操作方式使得用户无需具备专业的测量知识,就能够轻松完成测量任务。同时,仪器还配备了清晰的显示屏和直观的菜单,使得用户可以方便地查看测量结果和进行参数设置。 此外,土壤氧化还原电位检测仪还具有完善的数据记录和分析功能。仪器可以记录每次测量的结果,并提供相关的统计分析功能,方便用户对土壤氧化还原特性进行研究和评估。这种数据记录和分析功能使得用户能够更好地了解土壤状况的变化趋势,从而制定更为科学的农业生产计划和环境保护措施。 最后,土壤氧化还原电位检测仪还具有良好的耐用性和稳定性。它采用了高品质的材料和先进的制造工艺,确保了仪器的可靠性和稳定性。同时,仪器还具备防水、防尘等特性,可以在恶劣的环境条件下进行工作。这种耐用性和稳定性使得土壤氧化还原电位检测仪能够长期稳定地为用户提供服务。 综上所述,土壤氧化还原电位检测仪具有高精度测量、多功能、易操作性、数据记录和分析功能以及良好的耐用性和稳定性等众多优点。这些优点使得它在农业生产、环境保护以及科学实验等领域得到了广泛的应用,并为这些领域的发展提供了有力的支持。未来,随着科技的不断发展,土壤氧化还原电位检测仪的性能和功能还将得到进一步的提升和完善,为人类的生产和生活带来更多的便利和效益。
(一)标称阻值的检测 测量时,选用[URL=http://www.midiqi.com/Shop/Product.asp?ClassId=277]万用表[/URL] 绝缘万用表UT531 电阻档的适当量程,将两表笔分别接在[URL=http://www.midiqi.com/Shop/Product.asp?ClassId=363]电位器[/URL] SA-R10A研磨工具 两个固定引脚焊片之间,先测量电位器的总阻值是否与标称阻值相同。若测得的阻值为无穷大或较标称阻值大,则说明该电位器已开路或变值损坏。然后再将两表毛分别接电位器中心头与两个固定端中的任一端,慢慢转动电位器手柄,使其从一个极端位置旋转至另一个极端位置,正常的电位器,万用表表针指示的电阻值应从标称阻值(或0Ω)连续变化至0Ω(或标称阻值)。整个旋转过程中,表针应平稳变化,而不应有任何跳动现象。若在调节电阻值的过程中,表针有跳动现象,则说明该电位器存在接触不良的故障。 直滑式电位器的检测方法与此相同。 (二)带[URL=http://www.midiqi.com/Shop/Product.asp?ClassId=300]开关[/URL] 紧急停止开关HW 电位器的检测 对于带开关的电位器,除应按以上方法检测电位器的标称阻值及接触情况外,还应检测其开关是否正常。先[URL=http://www.midiqi.com/Shop/Product.asp?ClassId=369]旋转电位器[/URL] 旋转电位器R128 轴柄,检查开关是否灵活,接通、断开时是否有清脆的“喀哒”声。用万用表R×1Ω档,两表笔分别在电位器开关的两个外接焊片上,旋转电位器轴柄,使开关接通,万用表上指示的电阻值应由无穷大(∞)变为0Ω。再关断开关,万用表指针应从0Ω返回“∞”处。测量时应反复接通、断开电位器开关,观察开关每次动作的反应。若开关在“开”的位置阻值不为0Ω,在“关”的位置阻值不为无穷大,则说明该电位器的开关已损坏。 (三)双连同轴电位器的检测 用万用表电阻档的适当量程,分别测量双连电位器上两组电位器的电阻值(即A、C之间的电阻值和A’、C’之间的电阻值)是否相同且是否与标称阻值相符。再用导线分别将电位器A、C’及电位器A’、C短接,然后用万用表测量中心头B、B’之间的电阻值,在理想的情况下,无论电位器的转轴转到什么位置,B、B’两点之间的电阻值均应等于A、C或A’、C’两点之间的电阻值(即万用表指针应始终保持在A、C或A’、C’阻值的刻度上不动)。若万用表指针有偏转,则说明该电位器的同步性能不良。 图2-10是双连同轴电位器的电路图形符号。 [URL=http://www.midiqi.com/UploadFiles/Knowledge/20091125/200911250816461022.jpg]电路图形符号[/URL]
求教各位高手,钢结构自然腐蚀电位及保护电位用什么仪器检测,采用的规范是什么?肯请提供一份报告格式。
[b][url=http://www.f-lab.cn/microinjectors/explorer.html]游离脂质双层膜操作仪[/url][color=#666666][i],[/i][/color][i]Ionovation Explorer[/i][/b]把传统双分子层放置水平位置,自动生成[b]游离脂质双层[/b]并适合所有的标准倒置显微镜,包括从简单的光学显微镜到单分子光谱显微镜如FCS,FIDA,FRET等,游离脂质双层膜操作仪是生成[b]游离脂质双层分子膜[/b]进行分子膜研究的理想仪器,它客服了传统的垂直方法的局限,使得用户能够从生物膜两侧操作双层膜,改变介质和应用效应器等工作.游离脂质双层膜操作仪可作为显微镜配件,与高分辨率显微镜组成一流的单分子光谱技术电生理分析系统.它采用的紧凑结构设计适合任何倒置显微镜.提供自动膜生成,双侧灌注以及温度控制和监测功能。游离脂质双层膜操作仪配备了紧凑的“显微镜单元”和同样紧凑的“控制和泵单元”,也可以单独用作纯电生理实验系统。游离脂质双层膜操作仪与Explorer SLIDE同时使用时,可供了初学者和高级专业人员使用.[b][url=http://www.f-lab.cn/microinjectors/explorer.html]游离脂质双层膜操作仪[/url][/b]特色:重组的离子通道和毛孔从许多来源脂质翻转侧膜组织(脂筏)与离子通量直接相关的配体结合结合研究,根据预先设定的膜电位膜联蛋白聚合,聚合和跟踪在用户控制模型的各种膜的脂质成分人工膜上的拉曼光谱人工膜的探针应用[img=游离脂质双层膜操作仪]http://www.f-lab.cn/Upload/ionovation-explorer.jpg[/img]
检测气体的浓度依赖于气体检测变送器,传感器是其核心部分,按照检测原理的不同,主要分为金属氧化物半导体式传感器、催化燃烧式传感器、定电位电解式气体传感器、迦伐尼电池式氧气传感器、红外式传感器、PID光离子化传感器、等以下简单概述各种传感器的原理及特点。金属氧化物半导体式传感器金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。催化燃烧式传感器。催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。定电位电解式气体传感器定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。迦伐尼电池式氧气传感器隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。红外式传感器红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。PID光离子化气体传感器PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。
谁知道MOTT-Schottky时,为什么总是有这种现象比如以3V的电位递减到0V,曲线的大概在2.5V时斜率变为负的,2.5V以前斜率是正的.但是如果改变形成膜的电位2.0V极化1小时,再从2.0V递减到0V,这会还是快接近2.0V的时候斜率为负,之前的斜率为正 总是末尾的一小部分斜率为负,之前的斜率为正,是不是有问题
检测气体的浓度依赖于气体检测变送器,传感器是其核心部分,按照检测原理的不同,主要分为金属氧化物半导体式传感器、催化燃烧式传感器、定电位电解式气体传感器、迦伐尼电池式氧气传感器、红外式传感器、PID光离子化传感器、等以下简单概述各种传感器的原理及特点。金属氧化物半导体式传感器 金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。催化燃烧式传感器。 催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。定电位电解式气体传感器 定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。迦伐尼电池式氧气传感器 隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。红外式传感器 红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。PID光离子化气体传感器 PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。
传感器是气体检测变压器的核心部位,是检测气体浓度的关键所在,随着不同的检测原理,传感器也不尽相同。 PID光离子化气体传感器 PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。 迦伐尼电池式氧气传感器 隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。 催化燃烧式传感器 催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。www.jiuxing17.com 定电位电解式气体传感器 定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。 红外式传感器 红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。 金属氧化物半导体式传感器 金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。
简述几种气体检测传感器的检测原理此文章由 东方嘉仪仪器网 转发检测气体的浓度依赖于气体检测变送器,传感器是其核心部分,按照检测原理的不同,主要分为金属氧化物半导体式传感器、催化燃烧式传感器、定电位电解式气体传感器、迦伐尼电池式氧气传感器、红外式传感器、PID光离子化传感器等,以下简单阐述各种传感器的原理及特点。 金属氧化物半导体式传感器 金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。 催化燃烧式传感器 催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。 定电位电解式气体传感器 定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。 迦伐尼电池式氧气传感器 隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。 红外式传感器 红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。 PID光离子化气体传感器 PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。
检测气体的浓度依赖于气体检测变送器,传感器是其核心部分,按照检测原理的不同,主要分为金属氧化物半导体式传感器、催化燃烧式传感器、定电位电解式气体传感器、迦伐尼电池式氧气传感器、红外式传感器、PID光离子化传感器等,以下简单阐述各种传感器的原理及特点。 金属氧化物半导体式传感器 金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。 催化燃烧式传感器 催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。 定电位电解式气体传感器 定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。 迦伐尼电池式氧气传感器 隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。 红外式传感器 红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。 PID光离子化气体传感器 PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。
检测气体的浓度依赖于气体检测变送器,传感器是其核心部分,按照检测原理的不同,主要分为金属氧化物半导体式传感器、催化燃烧式传感器、定电位电解式气体传感器、迦伐尼电池式氧气传感器、红外式传感器、PID光离子化传感器等,以下简单阐述各种传感器的原理及特点。 金属氧化物半导体式传感器 金属氧化物半导体式传感器利用被测气体的吸附作用,改变半导体的电导率,通过电流变化的比较,激发报警电路。由于半导体式传感器测量时受环境影响较大,输出线形不稳定。金属氧化物半导体式传感器,因其反应十分灵敏,故目前广泛使用的领域为测量气体的微漏现象。 催化燃烧式传感器 催化燃烧式传感器原理是目前最广泛使用的检测可燃气体的原理之一,具有输出信号线形好、指数可靠、价格便宜、无与其他非可燃气体的交叉干扰等特点。催化燃烧式传感器采用惠斯通电桥原理,感应电阻与环境中的可燃气体发生无焰燃烧,使温度使感应电阻的阻值发生变化,打破电桥平衡,使之输出稳定的电流信号,再经过后期电路的放大、稳定和处理最终显示可靠的数值。 定电位电解式气体传感器 定电位电解式传感器是目前测毒类现场最广泛使用的一种技术,在此方面国外技术领先,因此此类传感器大都依赖进口。定电位电解式气体传感器的结构:在一个塑料制成的筒状池体内,安装工作电极、对电极和参比电极,在电极之间充满电解液,由多孔四氟乙烯做成的隔膜,在顶部封装。前置放大器与传感器电极的连接,在电极之间施加了一定的电位,使传感器处于工作状态。气体与的电解质内的工作电极发生氧化或还原反应,在对电极发生还原或氧化反应,电极的平衡电位发生变化,变化值与气体浓度成正比。 迦伐尼电池式氧气传感器 隔膜迦伐尼电池式氧气传感器的结构:在塑料容器的一面装有对氧气透过性良好的、厚10~30μm的聚四氟乙烯透气膜,在其容器内侧紧粘着贵金属(铂、黄金、银等)阴电极,在容器的另一面内侧或容器的空余部分形成阳极(用铅、镉等离子化倾向大的金属)。用氢氧化钾。氧气在通过电解质时在阴阳极发生氧化还原反应,使阳极金属离子化,释放出电子,电流的大小与氧气的多少成正比,由于整个反应中阳极金属有消耗,所以传感器需要定期更换。目前国内技术已日趋成熟,完全可以国产化此类传感器。 红外式传感器 红外式传感器利用各种元素对某个特定波长的吸收原理,具有抗中毒性好,反应灵敏,对大多数碳氢化合物都有反应。但结构复杂,成本高。 PID光离子化气体传感器 PID由紫外灯光源和离子室等主要部分构成,在离子室有正负电极,形成电场,待测气体在紫外灯的照射下,离子化,生成正负离子,在电极间形成电流,经放大输出信号。PID具有灵敏度高,无中毒问题,安全可靠等优点。 TOP







 我要推广仪器
我要推广仪器
 下载APP
下载APP