推荐厂家
暂无
暂无
[size=15px][font=宋体][color=black]内质网氨基肽酶[i][/i][/color][/font][font=&][color=black]1[/color][/font][font=宋体][color=black]([/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black])的主要功能是在内质网中修剪[/color][/font][font=&][color=black]N[/color][/font][font=宋体][color=black]端延长的肽前体,这是加工和呈递内源性抗原肽的关键步骤,它们被装载到[/color][/font][font=&][color=black]MHC-I[/color][/font][font=宋体][color=black]的凹槽中,在细胞表面呈递,激活[/color][/font][font=&][color=black]CD8+T[/color][/font][font=宋体][color=black]细胞或[/color][/font][font=&][color=black]NK[/color][/font][font=宋体][color=black]细胞,触发相应的免疫反应。[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]的过度活性加剧了相关的自身免疫疾病。此外,[/color][/font][font=&][color=black]ERAP1 [/color][/font][font=宋体][color=black]的过度活性会破坏肿瘤新抗原肽,导致肿瘤免疫逃逸。鉴于[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]在自身免疫疾病和肿瘤免疫逃逸中的关键作用,针对[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]功能的抑制将显著减少自身免疫疾病相关的疾病表型,同时也能抑制肿瘤免疫逃逸。因此,开发高效和选择性的[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]抑制剂已成为一个重要的药理学研究方向。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=宋体][color=black]通过高通量虚拟筛选结合物理筛选方法,从近[/color][/font][font=&][color=black]200000[/color][/font][font=宋体][color=black]种化合物中筛选到一种新的[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]选择性抑制剂[/color][/font][font=&][color=black]—[/color][/font][font=宋体][color=black]鼠尾草酸,并证明其与[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]有强烈的直接相互作用,通过竞争性抑制结合[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]的活性位点抑制其活性,进而抑制内质网应激,保持正常的抗原呈递功能。[/color][/font][font=&][color=black][/color][/font][/size] [align=center] [/align] [size=15px][b][font=&][color=#0070c0]1[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]ERAP1[/color][/font][font=宋体][color=#0070c0]抑制剂的筛选[/color][/font][/b][/size] [size=15px][font=宋体][color=black]作者首先通过虚拟筛选从[/color][/font][font=&][color=black]20[/color][/font][font=宋体][color=black]万个小分子化合物库[i][/i]中确定了对接排名靠前的[/color][/font][font=&][color=black]3250[/color][/font][font=宋体][color=black]个化合物,并定制了包含这些化合物的实体库用于酶活抑制效率的筛选,最终确定编号为“[/color][/font][font=&][color=black]3[/color][/font][font=微软雅黑, &][color=black]?[/color][/font][font=&][color=black]23[/color][/font][font=宋体][color=black]”的天然产物鼠尾草酸(广泛存在于唇科植物如迷迭香和鼠尾草)对[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]的酶活性抑制效果最佳。此外,鼠尾草酸对其他[/color][/font][font=&][color=black]6[/color][/font][font=宋体][color=black]种氨基肽酶均无明显抑制作用,通过竞争性抑制方式抑制[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]活性[/color][/font][/size][align=center] [/align] [size=15px][b][font=&][color=#0070c0]2[/color][/font][font=宋体][color=#0070c0]、鼠尾草酸能够直接与[/color][/font][font=&][color=#0070c0]ERAP1[/color][/font][font=宋体][color=#0070c0]结合形成稳定的配合物[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]接着,作者通过生物层干涉测量(BLI)[/color][/font][font=宋体][color=black]和细胞热移测定法(CETSA)[/color][/font][font=宋体][color=black]实验确定了鼠尾草酸与[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]直接结合,表明鼠尾草酸通过直接结合靶标[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black],进而抑制[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]活性 [/color][/font][/size] [size=15px][b][font=&][color=#0070c0]3[/color][/font][font=宋体][color=#0070c0]、鼠尾草酸与[/color][/font][font=&][color=#0070c0]ERAP1[/color][/font][font=宋体][color=#0070c0]的相互作用模式分析[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]进一步作者研究了鼠尾草酸与[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]的相互作用模式,通过“[/color][/font][font=&][color=black]ERAP1[/color][/font][font=微软雅黑, &][color=black]?[/color][/font][font=宋体][color=black]鼠尾草酸”对接配合物的最佳结构、相互作用能计算和分子动力学模拟等理论方法以及点突变的实验方法,发现“[/color][/font][font=&][color=black]ERAP1 -[/color][/font][font=宋体][color=black]鼠尾草酸”[/color][/font] [font=宋体][color=black]结合方式非常稳定,鼠尾草酸占据了[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]的催化中心,形成稳定的氢键网络,突变实验表明鼠尾草酸对[/color][/font][font=&][color=black]ERAP1 E183A[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]Q181A[/color][/font][font=宋体][color=black]突变体的抑制活性显著减少[/color][/font][font=宋体][color=black]。此外,鼠尾草酸与[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]ERAP2[/color][/font][font=宋体][color=black]结合模式存在显著不同 [/color][/font][/size] [size=15px][b][font=&][color=#0070c0]4[/color][/font][font=宋体][color=#0070c0]、鼠尾草酸逆转[/color][/font][font=&][color=#0070c0]ERAP1[/color][/font][font=宋体][color=#0070c0]引起的内质网应激反应[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]可通过过度剪切内源性抗原肽导致一系列免疫反应紊乱,如增加错误折叠[/color][/font][font=&][color=black]HLA[i][/i][/color][/font][font=宋体][color=black]分子组装的效率导致内质网应激。作者发现[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]过表达诱导内质网应激,体现在内质网应激标记蛋白[/color][/font][font=&][color=black]BIP[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]Chop[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]CANX[/color][/font][font=宋体][color=black]显著升高,而鼠尾草酸可抑制[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]引起的内质网应激[/color][/font][font=宋体][color=black]。[/color][/font][font=&][color=black][/color][/font][/size] [align=center][img=图片,1,]data:image/svg+xml,%3C%3Fxml version='1.0' encoding='UTF-8'%3F%3E%3Csvg width='1px' height='1px' viewBox='0 0 1 1' version='1.1' xmlns='http://www.w3.org/2000/svg' xmlns:xlink='http://www.w3.org/1999/xlink'%3E%3Ctitle%3E%3C/title%3E%3Cg stroke='none' stroke-width='1' fill='none' fill-rule='evenodd' fill-opacity='0'%3E%3Cg transform='translate(-249.000000, -126.000000)' fill='%23FFFFFF'%3E%3Crect x='249' y='126' width='1' height='1'%3E%3C/rect%3E%3C/g%3E%3C/g%3E%3C/svg%3E[/img][/align][align=center] [/align] [size=15px][b][font=&][color=#0070c0]5[/color][/font][font=宋体][color=#0070c0]、鼠尾草酸显著减少[/color][/font][font=&][color=#0070c0]ERAP1[/color][/font][font=宋体][color=#0070c0]引起的内源性抗原[i][/i]加工和递呈途径的中断[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]接着,作者评估了鼠尾草酸在细胞水平对抗原呈递功能的影响。[/color][/font][font=&][color=black]Western blot[/color][/font][font=宋体][color=black]和免疫荧光结果显示,[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]过表达导致内质网和细胞表面[/color][/font][font=&][color=black]HLA[/color][/font][font=宋体][color=black]水平升高,而鼠尾草酸的加入显著地扭转了这一趋势,减少细胞表面显示的额外大量[/color][/font][font=&][color=black]HLA[/color][/font][font=宋体][color=black]分子,以及减少细胞表面错误组装的[/color][/font][font=&][color=black]HLA[/color][/font][font=宋体][color=black]的比例,并保持正常的抗原呈递功能[/color][/font][/size] [align=center][img=图片,1,]data:image/svg+xml,%3C%3Fxml version='1.0' encoding='UTF-8'%3F%3E%3Csvg width='1px' height='1px' viewBox='0 0 1 1' version='1.1' xmlns='http://www.w3.org/2000/svg' xmlns:xlink='http://www.w3.org/1999/xlink'%3E%3Ctitle%3E%3C/title%3E%3Cg stroke='none' stroke-width='1' fill='none' fill-rule='evenodd' fill-opacity='0'%3E%3Cg transform='translate(-249.000000, -126.000000)' fill='%23FFFFFF'%3E%3Crect x='249' y='126' width='1' height='1'%3E%3C/rect%3E%3C/g%3E%3C/g%3E%3C/svg%3E[/img][/align][align=center] [/align] [size=15px][b][font=宋体][color=#0070c0]总结[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]该研究通过高通量虚拟筛选结合物理筛选方法,从近[/color][/font][font=&][color=black]200,000[/color][/font][font=宋体][color=black]种化合物中筛选出一种结构特异性化合物[/color][/font][font=&][color=black]—[/color][/font][font=宋体][color=black]鼠尾草酸。作者发现鼠尾草酸在蛋白质和细胞水平上与[/color][/font][font=&][color=black]ERAP1[/color][/font][font=宋体][color=black]有强烈的直接相互作用,通过竞争性抑制结合[/color][/font][font=&][color=black] ERAP1 [/color][/font][font=宋体][color=black]的活性位点,并且对同源蛋白[/color][/font][font=&][color=black] ERAP2 [/color][/font][font=宋体][color=black]以及广泛的相关代表性蛋白酶没有抑制活性,从而实现了有效的蛋白酶选择性[/color][/font][/size]
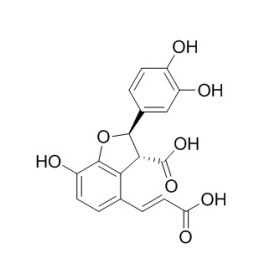
高效液相色普法测定甘草酸苷含量,甘草酸苷对照品峰面积与之前相比较偏高,导致含量偏高,请问有可能是哪些原因?
最近做小儿七星茶颗粒甘草酸的测定时遇到了很奇怪的问题:对照品有峰,样品却只出杂质峰,开始怀疑是样品没有含量,可是拿以前做过的有含量的样品再做,却没峰了;后来吸一半对照品一半样品进样就出峰了,可是加对照品到样品中一起按标准处理后就又没有峰出来,怎么想都不明白问题出在哪里。所用的试剂换了好几次,也换人配了,结果还是一样没有,会不会是超声引起的呢,因为我们的超声机的功率只有80瓦,不过以前也做得出啊,大家帮帮忙,下面是标准【含量测定】 照高效液相色谱法(中国药典2005年版一部附录VI D)测定。 色谱条件与系统适用性试验 以十八烷基硅烷键合硅胶为填充剂;以甲醇-0.2mol/L醋酸铵溶液—冰醋酸(65:35:1)为流动相;检测波长为250nm。理论板数按甘草酸峰计算应不低于2000。 对照品溶液的制备 取甘草酸铵对照品适量,精密称定,加流动相制成每1ml含16μg的溶液,即得(折合甘草酸为15.672μg)。 供试品溶液的制备 取装量差异项下的本品内容物,混匀,研细,取约7g,精密称定,置50ml量瓶中,加流动相约45ml,超声处理(功率300W,频率40kHz)30分钟,放冷,加流动相至刻度,摇匀,滤过,取续滤液,即得。 测定法 精密吸取对照品溶液与供试品溶液各20μl,注入液相色谱仪,测定,即得。







 400-659-9826
400-659-9826
 留言咨询
留言咨询
 留言咨询
留言咨询
 400-860-5168转3346
留言咨询
400-860-5168转3346
留言咨询









 我要推广仪器
我要推广仪器
 下载APP
下载APP