附件是原料巯基丙酮用酒精稀释后进的gcms,请问巯基丙酮二聚体的峰到底是14.866还是22.072,或者说两者都是?还有,根据香料通则,这个东西的含量要达到95%,根据图上看有个很大的巯基丙酮,含量应该不到95%,巯基丙酮是本来就有的呢还是二聚体分解出来的?大家做原料控制的时候怎么做的呢?
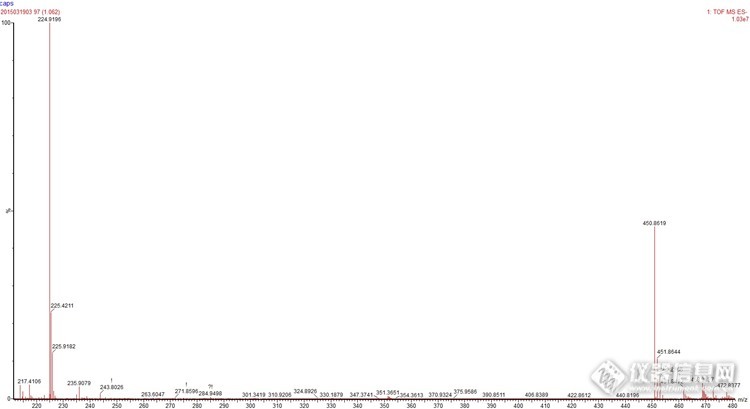
ES-做的,224.9同位素相差是0.5,450.9同位素相差1,这个是二聚体吗?分子量是452?大神解释下http://ng1.17img.cn/bbsfiles/images/2015/03/201503201055_538947_2359430_3.jpg
如题,乙偶因二聚体在气相上能出得来不?如何测定乙偶姻中二聚体的含量?
甲基环戊二烯二聚体有没有异构体?在[url=https://insevent.instrument.com.cn/t/Mp]气相[/url]上出几个峰?用什么色谱柱?
1.从引物自身着手,重新设计引物,这是最根本解决这一问题的办法。2.可能模板有问题,模板浓度过小,适当加大模板量。3.Taq酶,引物,Mg2+浓度可能过高,可降低它们的浓度。4.将上下引物混合后,在100℃的沸水中煮5分钟,然后迅速拿出至于冰块之上瞬时冷却,这时再加入反应体系当中,引物二聚体就会消失的。理由:引物可能会发生发夹结构,自身环化等结构,在100℃的沸水中煮5分钟可使引物变为单链,以减少二聚体。不过有人认为在PCR仪上95度变性5min也同样达到目的,而且成功试过通过延长退火时间也可以消除引物二聚体。5.所配MIX中加5%的甘油或者5%的DMSO,可以增强特异性。6.PCR反应体系的配制在冰上进行,最后加Taq酶,PCR结束后,产物勿放置在室温下过长时间,有人认为室温下有些Taq酶会将多余的引物合成为二聚体。7.增加循环数。8.降低退火温度后有条带,则应逐渐提高温度,若提高温度的同时产物量减少,则考虑增加Mg2+浓度(根据扩增片断长度而定,片段长则相应镁离子浓度应该高一些)。9.若降低退火温度,发现还是只有引物二聚体,而且镁离子的浓度在20-25mmol/l没有区别,则考虑Buffer等试剂没有完全融解、混匀,导致吸取的试剂浓度不对。10.以上次的PCR产物作模板二次PCR,可以提高引物与模板的特异性,减少引物二聚体,如果两次时间间隔短的话,可以把原产物稀释100-1000倍,如果间隔较长可以稀释50-100倍。
求二聚酸中单体、二聚体、三聚体含量测定方法?
二聚体的色谱校正因子是多少
我的化合物是属于含金属锂离子的复合物,然后又加了溴离子,(即得到了这样的二聚体,其他实验已证实该二聚体的存在),做了高分辨质谱发现了双电荷的峰,这个双电荷的 峰正好是显示含2个锂离子的二聚体的分子量那么我能不能判断我的东西在加了溴离子之后可以形成二聚体注:我的化合物不能络合两个金属锂离子,不加溴离子之前的高分辨质谱中也没发现二聚体的峰
在解析ESI低分辨时,如何区分有二倍关系的是二具体,还是因为出现了多点带电,比如说,我打了个ESI+低分辨300-500有个离子峰M/Z=415.2,响应强度为2.40e3。而600-1000范围有两个较强的离子峰M/Z=785.4,M/Z=807.4,响应强度为681。从785.4和807.4可判断出807.4为加Na,785.4为加H,415.2也是加Na。那么分子量到底是多少,按二聚体算的话,分子量应该为392,。如果说是415.2带了两个电荷,那么分子量是不是应该就是784。(请高手给讲讲,这ESI源打质谱如何判断分子量)图上传不上
请各位老师谈谈方向,我想试试GC测三氧化硫以及二聚体三聚体的含量。
[url=https://insevent.instrument.com.cn/t/Mp][color=#3333ff]气相[/color][/url]如何检测乙偶姻二聚体?查沸点279,出峰却很早,与乙偶姻出峰在一个位置上。
我想从猪肝中分离纯化金属硫蛋白,样品经过Sephadex G-75凝胶柱后,提出其中MT二聚体的蛋白峰,分子量14000左右,经过浓缩后上弱阴离子交换柱,采用0.01-1.0M醋酸铵缓冲液线性梯度洗脱,PH8.3,但是经过一次实验后溶液的PH降低,大量洗脱平衡后仍不能达到8.3,现在我该怎么办?请赐教.
我正在制备双特异抗体,采用半分子互换方法,即一半A 一半B,但是交联后,仍有少量的A和B的污染,因为我是想得到需要的双特异性抗体,因此需要纯化,请问如何才能做到? 据说可以用分析型CIEX实现 不知道具体的方法和原理如何 望指教,谢谢 我的联系方式时 13936179062 微信 电子邮件是 13936179062@139.com
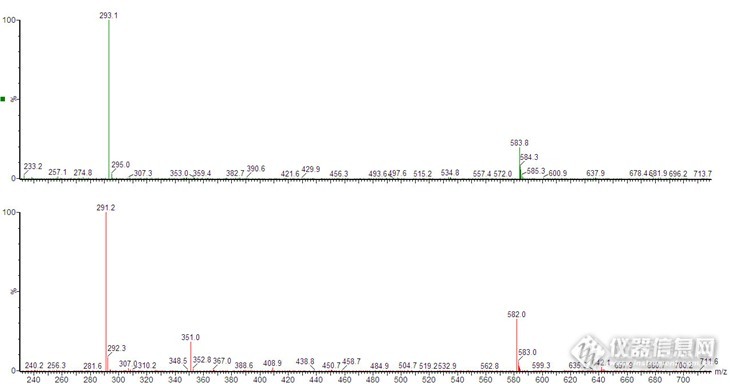
我有一个质谱图,已知分子离子峰在不同模式下分别为291和293,那么二聚体峰如何解释呢?见附图。data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAABPwAAAKfCAYAAAD6jdY4AAAgAElEQVR4AezdPVLb2v84/uP/ZAtQ3CY1A8VtswAYuuyBCRuAJt3nf7vbkA3AZA+3y8AC0n4LGGqaFGYR/kmyjy3JkiwbA/LhxUyip/P4OrIs3uhhNMl+gh8CBAgQIECAAAECBAgQIECAAAECBJIQ+P+S6IVOECBAgAABAgQIECBAgAABAgQIECBQCAj42REIECBAgAABAgQIECBAgAABAgQIJCQg4JfQYOoKAQIECBAgQIAAAQIECBAgQIAAAQE/+wABAgQIECBAgAABAgQIECBAgACBhAQE/BIaTF0hQIAAAQIECBAgQIAAAQIECBAgIOBnHyBAgAABAgQIECBAgAABAgQIECCQkICAX0KDqSsECBAgQIAAAQIECBAgQIAAAQIEBPzsAwQIECBAgAABAgQIECBAgAABAgQSEhDwS2gwdYUAAQIECBAgQIAAAQIECBAgQICAgJ99gAABAgQIECBAgAABAgQIECBAgEBCAgJ+CQ2mrhAgQIAAAQIECBAgQIAAAQIECBAQ8LMPECBAgAABAgQIECBAgAABAgQIEEhIQMAvocHUFQIECBAgQIAAAQIECBAgQIAAAQICfvYBAgQIECBAgAABAgQIECBAgAABAgkJCPglNJi6QoAAAQIECBAgQIAAAQIECBAgQEDAzz5AgAABAgQIECBAgAABAgQIECBAICEBAb+EBlNXCBAgQIAAAQIECBAgQIAAAQIECAj42QcIECBAgAABAgQIECBAgAABAgQIJCQg4JfQYOoKAQIECBAgQIAAAQIECBAgQIAAAQE/+wABAgQIECBAgAABAgQIECBAgACBhAQE/BIaTF0hQIAAAQIECBAgQIAAAQIECBAgIOBnHyBAgAABAgQIECBAgAABAgQIECCQkICAX0KD+SG78nwXLkejMJr9u3koKZS3Xd6F59Km8HAzzzMaXYa7ysZywtJ8Ud5NKFdR2mqWAAECBAgQIECAAAECBAgQIDAIAQG/QQyDRmwm8BBu9k/Cwf0kTCbZv/vrcH4Ug3fTbeF2XGwbn/4K+zEamAfujh6zTTHfQTjZXxXIm5b3Y7OGykWAAAECBAgQIECAAAECBAgQeDMBAb83o1bR1gUefofzi9vw9XBW8uHXcHvxIzyNs+XnP+ExXIez471i497xWbg+/zm9km/vOFxNrsJsUwiHX7KUj+FPy1V+z3eX2dWAR+Hx+jpcbL0TCiRAgAABAgQIECBAgAABAgQIbFdAwG+7nkp7S4HDb2FydRymIb284nF42uASvOe7n1ng8DT8vSio2ou/zsI4u4Lw6uvn6npLBAgQIECAAAECBAgQIECAAIEBCgj4DXBQNGkzgWngbnbF395f4SCch9+zB+4V28Ls6r9Y/OwZf9ldweH2ezlwGBNMp3uHh6WgYnWbJQIECBAgQIAAAQIECBAgQIDA0AQE/IY2ItqzkUB+2+3+yUG4n1/xdxi+jW/D49H0hR7/hrPsdt9a0cWtvfmz//o8w6+W1yIBAgQIECBAgAABAgQIECBAYKACAn4DHRjN6i8wDfZlV+mNv4X4OL8idwzo5bfjHofsdt+L8Hm/odwVz/BryGEVAQIECBAgQIAAAQIECBAgQGCwAp8G2zINI9BDYH5l36QW7KvnLV7icRC+tD2nr57eMgECBAgQIECAAAECBAgQIEBgRwV6XOH3HO4uR+Fm9iy0RT+n60ej6S2Tl3f1V5yu2r4oyRyBjQSyZ/D9mz9/r35lX1HYQ7gZXU7fypstP/x3En5cf5leAfhwk7119ybMd+k8GNj10o6NGicTAQIECBAgQIAAAQIECBAgQOB9BFYE/PKg3X44WXrz6Wz9wX2YZLdLTib34eBkPyyCfqu2v09n1ZqWQBHEy17EcbI/DTrH4PM0OJ09w694Nt9029HjbRh/m93wm73dd3z7GI5mwerR/lM4mz/7LwsO3oxK+3JaZnpDgAABAgQIECBAgAABAgQIpC8wygJ2k6ZuTm+VzCJ9F9fhOnvbaTibhBgvCfnbTfd/hdPxVTiOt0jmV039/BzGeeBk1famCq0jQIAAAQIECBAgQIAAAQIECBAgQODFAh1X+J2G+/zqvauv4XO9mvFT+FG/BXL/c7j48RTGedpV2+vlWSZAgAABAgQIECBAgAABAgQIECBAYCsCrQG/vePj6htPS9U9/3ksLZVnH8Of7FF+q7aXc5gnQIAAAQIECBAgQIAAAQIECBAgQGB7Aq0Bv+1V0VxSfN5a07Q5h7UEtisw+me03QKVRoAAAQIECBAgQIAAAQIECBAYgMCnTdqw99dBS7aD8Ff2TL9V2/PMTY8OzIN/fggQIECAAAECBAgQIECAAAECBAgQ2Fxgsyv8ys/ri3UXz+37HPbz5VXbYx5TAgQIECBAgAABAgQIECBAgAABAgS2KrBZwG/v73B6cR6Obh5mjXkIN0fn4eL071C8tHfV9q12QWEECBAgQIAAAQIECBAgQIAAAQIECESBzQJ+WVjv+Gocbh+PwvQZfEfh8XYcro6LcF9W9qrtsXpTAgQIECBAgAABAgQIECBAgAABAgS2KTDKnqU32WaBLykrPsNvQE16SXfkHbhA/tKOyf8Gs/sPXEvzCBAgQIAAAQIECBAgQIAAgV0R2PAKv13pnnYSIECAAAECBAgQIECAAAECBAgQ+FgCAn4fa7z1lgABAgQIECBAgAABAgQIECBAIHEBAb/EB1j3CBAgQIAAAQIECBAgQIAAAQIEPpaAgN/HGm+9JUCAAAECBAgQIECAAAECBAgQSFxAwC/xAdY9AgQIECBAgAABAgQIECBAgACBjyUg4PexxltvCRAgQIAAAQIECBAgQIAAAQIEEhcQ8Et8gHWPAAECBAgQIECAAAECBAgQIEDgYwkI+H2s8dZbAgQIECBAgAABAgQIECBAgACBxAUE/BIfYN0jQIAAAQIECBAgQIAAAQIECBD4WAICfh9rvPWWAAECBAgQIECAAAECBAgQIEAgcQEBv8QHWPcIECBAgAABAgQIECBAgAABAgQ+loCA38cab70lQIAAAQIECBAgQIAAAQIECBBIXEDAL/EB1j0CBAgQIECAAAECBAgQIECAAIGPJSDg97HGW28JECBAgAABAgQIECBAgAABAgQSFxDwS3yAdY8AAQIECBAgQIAAAQIECBAgQOBjCQj4fazx1lsCBAgQIECAAAECBAgQIECAAIHEBQT8Eh9g3SNAgAABAgQIECBAgAABAgQIEPhYAgJ+H2u89ZYAAQIECBAgQIAAAQIECBAgQCBxAQG/xAdY9wgQIECAAAECBAgQIECAAAECBD6WgIDfxxpvvSVAgAABAgQIECBAgAABAgQIEEhcQMAv8QHWPQIECBAgQIAAAQIECBAgQIAAgY8lIOD3scZbbwkQIECAAAECBAgQIECAAAECBBIX+DSE/o1GoyE0QxsIECBAgAABAgQIECBAgAABAgQI7LzAIK7wm0wmIf/nhwABAgQIECBAgAABAgQIECBAgACBlwkMIuD3si7ITYAAAQIECBAgQIAAAQIECBAgQIBAFBDwixKmBAgQIECAAAECBAgQIECAAAECBBIQEPBLYBB1gQABAgQIECBAgAABAgQIECBAgEAUEPCLEqYECBAgQIAAAQIECBAgQIAAAQIEEhAQ8EtgEHWBAAECBAgQIECAAAECBAgQIECAQBQQ8IsSpgQIECBAgAABAgQIECBAgAABAgQSEBDwS2AQdYEAAQIECBAgQIAAAQIECBAgQIBAFBDwixKmBAgQIECAAAECBAgQIECAAAECBBIQEPBLYBB1gQABAgQIECBAgAABAgQIECBAgEAUEPCLEqYECBAgQIAAAQIECBAgQIAAAQIEEhAQ8EtgEHWBA
多方求助无果,希望有高人指点。急急我们实验室在做重组人复合α干扰素(cIFN)的聚合与降解研究。cIFN单体中有两条分子内二硫键,它们在巯基乙醇作用下可以被还原打开,并且在空气中会形成分子间二硫键,进而引起cIFN聚合。再向这些聚合体中添加过量巯基乙醇后绝大多数的聚合体都被离解成单体,但是通过还原SDS-PAGE仍可以看到有微量的二聚体。理论上还原条件下二硫键是不存在的,所以我们推测这个二聚体应该是其他化学键引起的。我想知道的是:1、蛋白质除了二硫键是否还有其他的化学键可以引起蛋白质共价聚合体。2、用什么方法可以鉴定蛋白质聚合体中单体间的化学键?[em0812][em0812][em0811][em0811][em0811]
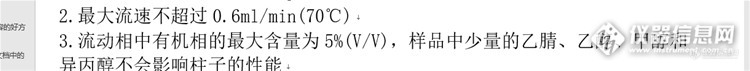
http://ng1.17img.cn/bbsfiles/images/2016/01/201601032253_580610_1621538_3.jpghttp://ng1.17img.cn/bbsfiles/images/2016/01/201601032253_580611_1621538_3.pnghttp://ng1.17img.cn/bbsfiles/images/2016/01/201601032253_580612_1621538_3.jpg帮一个同学问的:自己合成的样品,用液相色谱法分离,总是分不开。一个大峰,峰形不太好看,件图1.色谱柱:sugar-Ca;色谱柱要求见图2。分析标准品可以分开,见图3.现在流动相为水和乙醇,流速和柱温都调整了,还是分不开,请大家指点。
人血白蛋白多聚体测定法本法系用分子排阻色谱法测定人血白蛋白多聚体含量。照分子排阻色谱法(附录Ⅲ D)测定。色谱条件与系统适用性试验 用亲水硅胶高效体积排阻色谱柱(SEC,排阻极限300kD,粒度10μm),柱直径7.5mm,长60cm;以含1%异丙醇的pH7.0 0.2mol/L磷酸盐缓冲液为流动相;检测波长为280nm ;流速为每分钟0.6ml。取每1ml含蛋白质为12mg 的人血白蛋白溶液20μl,注入色谱柱,记录色谱图,人血白蛋白单体峰与二聚体峰间的分离度应大于1.5,拖尾因子按人血白蛋白单体峰计算应为0.95~1.40。测定法 取供试品适量,用流动相稀释成每1ml约含蛋白质12mg的溶液,取20μl,注入色谱柱,记录色谱图60分钟(色谱柱长60cm)。按面积归一法计算,色谱图中未保留(全排阻)峰的含量(%)除以2,即为人血白蛋白多聚体含量请教:为何除以2呢?很奇怪呀
我是学超分子化学的,以前我用NMR稀释的方法,通过记录特征H化学位移的变化,应用Origin进行非线性拟和可以求得氢键二聚体的结合常数。但对于结合常数高的二聚体,由于NMR灵敏度的限制,在浓度很小的情况下难以进行测定。现打算采用UV-vis(适宜低浓度)来进行测定,但我对UV-vis很陌生,特向大家请教!
在做二羟基丙酮的硅烷化分析的时候,出现一个问题:当标准样品的量比较少的时候比如10mg左右的时候,气相上出来的是二羟基丙酮的硅烷化的峰,但是当标品的来那个超过20mg的时候会出来二羟基丙酮硅烷化峰以及二羟基丙酮二聚体硅烷化峰,不知哪位专家做过此物质的分析,可否指点一下。
跟大家分享一下,聚氨酯的IR分析 v3250-3500 ms OH伸缩振动、NHCO的顺式NH伸缩振动。 v2940、2860 s CH2、CH3伸缩振动。 v2240-2280 s NCO特征吸收峰。 v2120 s 碳化二亚胺吸收峰。 v1770-1785 s 脲二酮环(二聚体)中的C=O。 v1715-1750 vs 酯基C=O、酰胺I键C=O。 v1689-1710 s 异腈脲酸酯(三聚体)中C=O(1408-1430也有峰) v1600-1615 苯环C=C骨架伸缩振动。 v1520-1560 ms 酰胺II键(N-H)变形振动。 v1450-1470 CH2变形振动、CH3非对称变形振动。 v1380 CH3对称变形振动 v1225-1235 聚酯C-O伸缩或OH变形振动 v1060-1150 宽s C-O-C(脂肪族醚)吸收峰。
在检测多聚体含量的时候,跟半个月前做的样品检测的单体峰的峰高以及峰面积都都有明显的下降,但是含量的比例还是保持稳定的。单体峰的峰高之前可以达到350-500mV都有的,基本稳定在400mV左右,但是今天做出来的单体峰的峰高却只有160mV,而且多聚体和二聚体也是一样的降了近一半,还有流动相是一样的,都是磷酸盐,柱子是去年7月份买的,请问下这个原因可能有哪些方面造成的?
我做的合成实验由于容易生成二聚体、多聚体,一般用GPC来大概地看下合成产物中的单体含量,但是合成单体为三种同分异构体的混合物,我想请教:1、同分异构体在GPC中会表现出差别吗?(分子量虽然相同,但是结构不同的话流体力学体积应该不同吧,有影响吗?)2、有差别的话,表现在什么地方呢?请高手赐教,多谢!
1.O-H伸缩振动羧酸在浓溶液或固态中因强氢键成二聚体。 强氢键源于离子共振,阻碍游离羟基振动,仅稀非极性溶剂或蒸气相中可见(约3520 cm?1)。 二聚体O-H振动宽且强,范围3300~2500 cm?1,常集中于3000 cm?1,伴弱C-H振动。 长波长精细结构为倍频与复合频。 β-二酮等也有此吸收,但较弱,C-O振动频率较低。与醚类溶剂形成分子间氢键,O-H吸收约3100 cm?1。 2.C-O伸缩振动羧酸C-O振动强于酮,单体约1760 cm?1。二聚体对称,仅不对称振动有吸收,氢键与共振降低频率至1720~1706 cm?1。 分子内氢键影响更大,如水杨酸1665 cm?1,对羟基苯甲酸1680 cm?1。 不饱和共轭轻微降低频率,α,β-不饱和及芳基共轭酸二聚体约1710~1680 cm?1。 α位电负性取代基(如卤素)轻微增加频率,旋转异构致双重谱带。 3.C-O伸缩与O-H弯曲振动羧酸红外光谱中,C-O伸缩约13201210 cm?1,O-H弯曲约14401395 cm?1,两者有相互作用。 二聚体C-O伸缩强吸收约1315~1280 cm?1,长链脂肪酸呈双峰。 O-H面外弯曲特征谱带约920 cm?1,中等强度峰宽。
聚丙烯腈纤维中加入第二、第三单体的作用答:丙烯腈为第一单体,它是聚丙烯腈纤维的主体,对纤维的许多化学、物理及力学性能起着主要的作用。第二单体为结构单体,加入量为5%~10%,通常选用含酯基的乙烯基单体,这些单体的取代基极性较氰基弱,基团体积又不大,可以减弱聚丙烯腈大分子间的作用力,从而改善纤维的手感和弹性,克服纤维的脆性,也有利于染料分子进人纤维内部。第三单体又称染色单体,是使纤维引入具有染色性能的基团,改善纤维的染色性能,一般选用可离子化乙烯基单体,加入量为0.5%~3%。
[align=center][b][font=宋体]附录Ⅵ[/font][font=Times New Roman] Q[/font][font=宋体]人血白蛋白多聚体测定法[/font][/b][/align][font=宋体][size=3]本法系用分子排阻色谱法测定人血白蛋白多聚体含量。[/size][/font][size=3][font=宋体]照分子排阻色谱法(附录Ⅲ[/font][font=Times New Roman] D[/font][font=宋体])测定。[/font][/size][size=3][b][font=宋体]色谱条件与系统适用性试验[/font][/b][font=Times New Roman] [/font][font=宋体]用亲水硅胶高效体积排阻色谱柱([/font][font=Times New Roman]SEC[/font][font=宋体],排阻极限[/font][font=Times New Roman]300kD[/font][font=宋体],粒度[/font][font=Times New Roman]10[/font][font=宋体]μ[/font][font=Times New Roman]m[/font][font=宋体]),柱直径[/font][font=Times New Roman]7[/font][font=宋体].[/font][font=Times New Roman]5mm[/font][font=Times New Roman],[/font][font=宋体]长[/font][font=Times New Roman]60cm[/font][font=宋体];以含[/font][font=Times New Roman]1[/font][font=宋体]%异丙醇的[/font][font=Times New Roman]pH7[/font][font=宋体].[/font][font=Times New Roman]0 0[/font][font=宋体].[/font][font=Times New Roman]2mol/L[/font][font=宋体]磷酸盐缓冲液[取[/font][font=Times New Roman]0[/font][font=宋体].[/font][font=Times New Roman]5mol/L[/font][font=宋体]磷酸二氢钠[/font][font=Times New Roman]200ml[/font][font=宋体]、[/font][font=Times New Roman]0[/font][font=宋体].[/font][font=Times New Roman]5mol/L[/font][font=宋体]磷酸氢二钠[/font][font=Times New Roman]420ml[/font][font=宋体]、异丙醇[/font][font=Times New Roman]15[/font][font=宋体].[/font][font=Times New Roman]5ml[/font][font=宋体]及水[/font][font=Times New Roman]914.5ml[/font][font=宋体],混匀]为流动相;检测波长为[/font][font=Times New Roman]280nm [/font][font=宋体];流速为每分钟[/font][font=Times New Roman]0[/font][font=宋体].[/font][font=Times New Roman]6ml[/font][font=宋体]。取每[/font][font=Times New Roman]1ml[/font][font=宋体]含蛋白质为[/font][font=Times New Roman]12mg [/font][font=宋体]的人血白蛋白溶液[/font][font=Times New Roman]20[/font][font=宋体]μ[/font][font=Times New Roman]l[/font][font=宋体],注入色谱柱,记录色谱图[/font][font=Times New Roman],[/font][font=宋体]人血白蛋白单体峰与二聚体峰间的分离度应大于[/font][font=Times New Roman]1[/font][font=宋体].[/font][font=Times New Roman]5[/font][font=宋体],拖尾因子按人血白蛋白单体峰计算应为[/font][font=Times New Roman]0[/font][font=宋体].[/font][font=Times New Roman]95[/font][font=宋体]~[/font][font=Times New Roman]1[/font][font=宋体].[/font][font=Times New Roman]40[/font][font=宋体]。[/font][/size][size=3][b][font=宋体]测定法[/font][/b][font=Times New Roman] [/font][font=宋体]取供试品适量,用流动相稀释成每[/font][font=Times New Roman]1ml[/font][font=宋体]约含蛋白质[/font][font=Times New Roman]12mg[/font][font=宋体]的溶液[/font][font=Times New Roman],[/font][font=宋体]取[/font][font=Times New Roman]20[/font][font=宋体]μ[/font][font=Times New Roman]l[/font][font=宋体],注入色谱柱,记录色谱图[/font][font=Times New Roman]60[/font][font=宋体]分钟[/font][s][font=宋体](色谱柱长[/font][/s][s][font=Times New Roman]60cm[/font][/s][s][font=宋体])[/font][/s][font=宋体]。[/font][/size][size=3][color=#fe2419][font=宋体]按面积归一法计算,色谱图中未保留(全排阻)峰的含量[/font][font=Times New Roman](%)[/font][font=宋体]除以[/font][font=Times New Roman]2[/font][font=宋体],即为人血白蛋白多聚体含量。[/font][/color][/size][size=3][font=Times New Roman][b][color=#0021b0]计算方法不是很明白,为什么要除以2 ??? 全排阻的峰指的是哪一个??谢谢[/color][/b][/font][/size]
我们现在要分析乙二醇,想自己做聚乙二醇20000填充柱,不知担体用什么?高手请指教一下,谢谢!
二甲基聚硅氧烷混合环体色谱法怎么做?本品为易燃物,生成碳氧化合物和二氧化硅。缺氧条件加热至200到300度时,分解生成甲醛,氧化剂能引起反应,
岛津HPLC(型号10A)在测定白蛋白多聚体的时应新药典要求进行系统适应性试验之分离度的计算class-LC10工作站中提供了直接计算的功能,但此项功能多数情况下不能给出二聚体峰与单体峰之间的分离度R,观察多张图谱得出只要是两峰中任一峰无理论塔板数则R无数值显示,即使是两峰在图谱上可明显观察到已分离,不明白为什么会出现此情况。PS:测定白蛋白成品时分析时间为70min,在单体峰(28min)之后会出一乙酰色氨酸峰(50min)而此两峰间R可达16左右。成品之前有一组分Ⅴ(生产中间体)样品也会测定其中多聚体含量,因其内不含乙酰色氨酸,故分析时间为50min,奇怪的就是恰恰因为这个原因使得每次单体峰与二聚体峰均可达到药典要求(R2.5)其实组分Ⅴ与成品组成几乎相同,仅仅只是纯度不及后者本人实在是不明白既然R是有公式计算得的,有峰存在,即有公式中的参数值即使是R很下,但也该有个数值吧!请高手帮忙解决一下,不胜感激!~
[size=14px] [/size] [size=14px]心肌梗死性心力衰竭(HF)是世界范围内导致死亡和残疾的主要原因,心衰的特征是线粒体能量产生受损,类视黄醇X受体a(RXRa)是一种配体激活的转录因子,可以形成同型二聚体或异源二聚体,激活参与底物利用和氧化磷酸化的基因转录,是调节线粒体能量代谢的理想靶点。然而,目前缺乏具有高亲和力和特异性激活RXRa二聚体的激动剂。[/size] [size=14px]黄芪甲苷是黄芪的主要有效成分,已有研究报道多种黄芪甲苷可以改善线粒体功能。其中黄芪甲苷Ⅳ已被观察到在腹膜纤维化中调节RXRa,这意味着黄芪甲苷家族的化合物具有激活RXRa和改善线粒体功能的潜力。因此,作者推测黄芪甲苷可能作为RXRa同型二聚体的潜在激动剂。此外,作者还研究了靶向RXRa的下游机制。[/size] [size=14px] [/size] [size=14px]黄芪甲苷III(AS-III)是一种理想的直接靶向RXRa的激动剂。机制上,AS-III通过与RXRa相互作用激活RXRa二聚体,随后上调Ndufs4的转录。更重要的是,AS-III在体外和体内均有助于改善线粒体损伤和心功能障碍。该研究为心衰治疗提供了一种新的治疗策略,通过RXRa同型二聚体的反激活来上调Ndufs4并促进能量代谢。[/size] [size=14px]图片[/size] [size=14px]1、AS-III靶向RXRa,保护线粒体功能,发挥心脏保护作用[/size] [size=14px]作者首先通过通过分子对接、表面等离子体共振(SPR)和微尺度热电泳(MST),鉴定出黄芪甲苷III(AS-III)是黄芪甲苷家族中亲和力最高的化合物(附件中)。SPR(图1a)和MST(图1b)结果显示AS-III特异性结合RXRa, KD值分别为3.05x10^-7和1.35x10^-7 mol/L,表明AS-III对RXR具有较高的亲和力。分子对接显示AS-III成功地停靠在二聚体的界面上(图1c)。对AS-III和RXRa蛋白进行均方根偏差(RMSD)模拟,结果表明它们在20 ns后达到稳定构象(图1d)。由于Ccl6受RXRa同型二聚体的转录调控,作者使用Ccl6启动子荧光素酶报告基因发现AS-III成功诱导了Ccl6的转录,表明AS-III是一种具有高亲和力的RXRa激动剂(附件)。采用体外氧葡萄糖剥夺/恢复(OGD/R)模型和体内左前降(LAD)结扎诱导HF模型确定了AS-III的保护作用,其作用与临床治疗HF的药物辛伐他汀相似(图1f和1g)。RXRa siRNA一致地逆转了AS-III依赖性的OGD/R损伤的保护作用,表明RXRa是AS-III的关键治疗靶点(图1e)。AS-III低剂量和高剂量的心脏保护作用相当,因此后续实验选择低剂量AS-III。随后采用透射电镜(TEM)和高效[url=https://insevent.instrument.com.cn/t/5p][color=#3333ff]液相色谱[/color][/url](HPLC)检测心肌组织线粒体结构和功能,发现AS-III部分减轻线粒体损伤(图1h),AS-III改善了ATP浓度(图1i),表明它促进了HF小鼠ATP的产生。此外,western blotting显示AS-III在小鼠HF模型中抑制RXRa下调(图1j)。RNA测序分析结合q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]验证证实,AS-III治疗上调了线粒体复合物亚基(Ndufs1-6和8),它们在模型组中严重受损(图1k和1)。[/size] [size=14px] [/size] [size=14px] [/size] [size=14px] [/size] [size=14px]2、AS-III通过激活RXRa上调Ndufs4在心脏中的表达促进线粒体功能[/size] [size=14px]越来越多的证据表明RXRa以依赖配体激活的方式调节线粒体复合体I,但其调控机制尚不明确。结合现有文献和前面的实验结果,作者推测RXRa通过Ndufs调控线粒体复合体I。作者利用HUMAN TFDB网站和Cytoscape软件确定Ndufs1-8是RXRa的直接转录靶基因。接着利用q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]验证RXRa与Ndufs1-8之间的关系,发现RXRa siRNA下调Ndufs2、Ndufs4、Ndufs6和Ndufs8(图2a),而RXRa过表达上调了Ndufs4(图2b)。Western blot结果显示RXRa敲低显著降低了Ndufs4水平,RXRa过表达上调了Ndufs4水平(附件)。CUT和Tagseq分析用于确定RXRa和nduf4之间的调控关系,在Ndufs4基因中鉴定出RXRa结合序列(图2c)。此外,采用ChIP-q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]方法证实RXRa与Ndufs4的互作。双荧光素酶报告基因实验发现RXRa激活了ndufs4驱动的荧光素酶基因表达。然而,在突变的Ndufs4荧光素酶活性中没有观察到这种差异(图2d)。这些结果支持RXRa直接靶向Ndufs4从而促进其转录的观点。q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]结果显示在转染RXRa质粒的H9c2细胞中,9-cis RA(RXRa泛激动剂)能诱导Ndufs4表达,而LG100754(PPAR/RXR的激动剂,但是RXR同型二聚体的拮抗剂)不能诱导Ndufs4表达,且LG100754和9-cis RA共处理未能上调Ndufs4的表达,这表明RXRa以其同二聚体形式调控Ndufs4的转录(图2e)。进一步使用已知与RXRa同型二聚体结合的rxre -荧光素酶报告基因发现AS-III通过激活RXRa同型二聚体来发挥作用。与分子对接结果一致,AS-III可以诱导RXRa同型二聚体的转录激活(图2f)。此外,AS-III上调了Ndufs4 mRNA的表达,RXRa siRNA显著减弱了Ndufs4 mRNA的表达(图2g)。还有,AS-III改善了OGD/R损伤的线粒体结构和功能,在LAD模型中发现AS-III通过rxra - ndufs4介导的线粒体功能发挥心脏保护作用(图2h,i)。Western blot结果显示AS-III处理强烈诱导Ndufs4表达,然而这种效应在AAV-RXRa KD小鼠中被破坏(2j)。在AAV-RXRa KD + LAD结扎组,AS-III对线粒体的保护作用减弱(图2K)。[/size]
样品是N-羟甲基丙烯酰胺,化学式是C4H7NO2。如何根据ESI质谱图分析是二聚体还是三聚体?谢谢了!







 我要推广仪器
我要推广仪器
 下载APP
下载APP