老板再也不用担心我的多肽合成 ---来阿拉丁一站式购齐所需试剂和容器
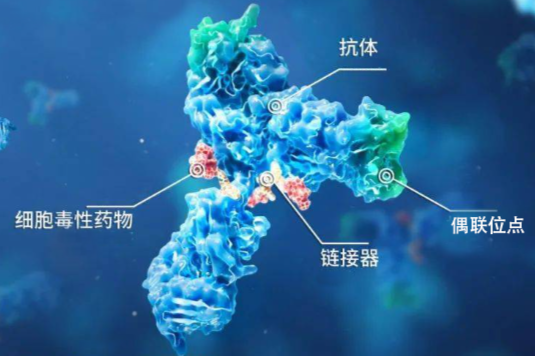
ALADDIN的优势多肽在基础生理学、生物化学和医药研究,尤其是医药行业新药筛选中起关键作用,新的短链肽和模拟肽在新药研发中为新药提供了较强的生物活性和蛋白酶水解抗性。短肽还可以作为分子探针,更好的阐述生物系统的功能。因此肽合成在化学生物学领域所占份额越来越大。阿拉丁为你提供高质固相和液相肽合成的一站式服务,包括带有Fmoc、Boc和Cbz保护基团的天然或非天然氨基酸合成砌块、偶联试剂、预装树脂、Linker、N-保护试剂。产品列表多肽固相合成管固相多肽合成预装树脂N-保护试剂耦合试剂Fmoc修饰的氨基酸及氨基酸衍生物列表Boc修饰的氨基酸及氨基酸衍生物列表更多相关产品耗材产品列表多肽固相合成管货号品名包装容量外径螺纹口砂板孔隙度P3597-01-1EAP3597-01 多肽固相合成管1个25ml25mm25G2P3597-02-1EAP3597-02 多肽固相合成管1个25ml25mm25G3 试剂产品列表固相多肽合成预装树脂货号品名规格包装 A116077Fmoc-Arg(Pbf)-Wang resin100-200 mesh, 1%DVB1g,5g,25g A116080Fmoc-Asn(Trt)-王氏树脂 100-200 mesh, 1%DVB,Substitution 0.41g,5g,25g A116082Fmoc-Asp(OtBu)-王氏树脂100-200 mesh, 1%DVB,Substitution 0.1g,5g,25g A118255Fmoc-氨基酸-王树脂100-200 mesh, 1%DVB,Substitution 0.3-0.8mmol/g5g,25g A118270AminoMethyl Polystyrene Resin0.5~1.5mmol/g, 100~200 mesh5g,25g,100g C110262氯甲基化聚苯乙烯树脂1% DVB交联 1.0~1.24mmol/g , 100~200 mesh, 1% DVB5g,25g,100g C1182692-Chlorotrityl Chloride Resin0.8-1.5mmol/g, 100~200 mesh5g,25g,100g G116092Fmoc-Glu(OtBu)-王氏树脂 100-200 mesh, 1%DVB,Substitution 0.1g,5g G116094Fmoc-Gly-Wang resin100-200 mesh, Substitution 0.3-0.8mmol/g5g,25g L116104Fmoc-Leu-王氏树脂100-200 mesh, Substitution 0.3-0.8mmol/g5g,25g L116107Fmoc-Lys(Boc)-王氏树脂 100-200 mesh, 1%DVB,Substitution 0.3-1g,5g,25g M118256Fmoc-Met-王氏树脂100-200 mesh, 1%DVB,Substitution 0.3-0.1g,5g,25g M118275MBHA Resin0.3~0.8mmol/g, 100~200 mesh, 1% DVB1g,5g,25g P118257Fmoc-D-Phe-王氏树脂 100-200 mesh, 1%DVB,Substitution 0.3-0.5g,25g P118258Fmoc-Phe(4-Cl)-Wang resin100-200 mesh, 1%DVB1g,5g,25g P118261Fmoc-Pro-王氏树脂 100-200 mesh, 1%DVB,Substitution 0.3-0.8m5g,25g R118279Rink Amide-AM Resin 0.3~0.8mmol/g, 100~200 mesh, 1% DVB1g,5g,25g R118280聚合物键合型 Rink 酰胺 4-甲基二苯甲胺0.3~0.8mmol/g, 100~2001g,5g,25g S118282Sieber 酰胺树脂0.3~0.8mmol/g, 100~200 mesh, 1% DVB5g,25g,100g T118264Fmoc-Thr(tBu)-王氏树脂100-200 mesh, 1%DVB,Substitution 0.31g,5g,25g T118267Fmoc-Tyr(tBu)-Wang resin100-200 mesh, 1%DVB,Substitution 0.5g,25g T118281Fmoc-Threoninol(tBu) DHP HM Resin 0.3~0.8mmol/g, 100~200 mes5g,25g V118268Fmoc-Val-Wang resin100-200 mesh, 1%DVB,Substitution 0.3-0.85g,25gN-保护试剂氨基保护是合成化学和肽合成中必须部分,有效的保护基团可以从合成的化合物易于添加和除去。货号品名规格cas号包装 B105737氯甲酸苄酯 96%,含约 0.1% 碳酸钠稳定剂501-53-125g,100g,500g,2.5kg D106158二碳酸二叔丁酯 98%24424-99-525g,100g,500g,1kg D106159二碳酸二叔丁酯 99%24424-99-525g,100g,1kg D106160二碳酸二叔丁酯 96%24424-99-5100g,500g F1061739-芴甲基-N-琥珀酰亚胺基碳酸酯 98%82911-69-15g,25g,100g F113338芴甲氧羰酰胺 99%84418-43-95g,25g,100g I105738氯甲酸异丁酯 98%543-27-125g,100g,500g耦合试剂由于肽合成中较低的消旋化是固相肽合成的一个关键指标,阿拉丁为你提供各种高质量偶联试剂,包括碳化二亚胺、脲类和磷型的偶联试剂,可以快速、有效和无消旋的缩合货号品名规格cas号包装 A1133452-(7-氮杂苯并三氮唑)-N,N,N' ,N' -四甲基脲四氟硼酸盐 98%873798-09-55g,25g,100g B106161卡特缩合剂 98%56602-33-65g,25g,100g,500g B1093122-溴-1-乙基吡啶四氟硼酸盐 98%878-23-95g,25g B113336溴代三(二甲基氨基)磷鎓六氟磷酸盐 98%50296-37-21g,5g,25g B113343三吡咯烷基溴化鏻六氟磷酸盐 98%132705-51-21g C109314N,N' -羰基二咪唑 &ge 97.0% (T)530-62-12.5kg,25g,100g,500g C109315N,N' -羰基二咪唑 99%530-62-11kg C113337N,N' -羰基二(1,2,4-三氮唑) 96%41864-22-65g,25g,100g H1061761-羟基苯并三唑一水合物 &ge 97.0%123333-53-925g,100g,250g,500g H1061773-羟基-1,2,3-苯并三嗪-4(3H)-酮 98%28230-32-25g,25g,100g H106354N-羟基邻苯二甲酰亚胺 98%524-38-92.5kg,25g,100g,500g H1093281-羟基-7-偶氮苯并三氮唑 99%39968-33-75g,25g,100g,500g H109329N-羟基-5-降冰片稀-2,3-二酰亚胺 99%21715-90-210g,50g,250gH109330N-羟基琥珀酰亚胺 98%6066-82-62.5kg,25g,100g,500g H109337N-羟基硫代琥珀酰亚胺 钠盐 98%106627-54-71g,5g,25g N102772N-琥珀酰亚胺基-N-甲基氨基甲酸酯 97%18342-66-05g,25g N113351TNTU 98%125700-73-41g,5g,25g,100g C113347多肽试剂TCTU 98%330641-16-25g,25g,100g C1171602-氯-1,3-二甲基咪唑六氟磷酸盐 98%101385-69-71g,5g,25g D1028482-(2-吡啶酮-1-基)-1,1,3,3-四甲基脲四氟硼酸盐 99%125700-71-21g,5g,25g D106162N,N' -二异丙基碳二酰亚胺(DIC) 98%693-13-010ml,25ml,100ml,500ml D106171N,N' -琥珀酰亚胺基碳酸酯 98%74124-79-15g,25g,100g D106284N,N-二甲基丙烯基脲(DMPU) 99%7226-23-525g,100g,500g D109331二吡咯烷基(N-琥珀酰亚氨氧基)碳六氟磷酸盐 98%207683-26-91g,5g,25g O113352TOTT 98%255825-38-85g,25g,100g P1091051-苯基-3-甲基-5-吡唑啉酮 99%89-25-82.5kg,100g,500g W111795伍德沃德氏试剂K 98%4156-16-51gFmoc修饰的氨基酸及氨基酸衍生物列表货号品名规格cas号包装 A107817Fmoc-L-天冬氨酸 4-烯丙酯 98%146982-24-31g,5g,25g A140203N-Fmoc-8-氨基辛酸 &ge 98.0%(HPLC)126631-93-41g,5g B116715N-Boc-N' -Fmoc-D-赖氨酸 97%115186-31-75g,25g B121679N-Boc-顺式-4-Fmoc-氨基-L-脯氨酸 97%174148-03-91g,5g C115874FMOC-&beta -环己基-L-丙氨酸 98%135673-97-11g,5g,25g C115932Fmoc-Cys(Mbzl)-OH 98%136050-67-41g,5g,25g D115880N&alpha -Fmoc-L-2,3-二氨基丙酸 97%181954-34-71g,5g,25g F100409Fmoc-S-三苯甲基-L-半胱氨酸 98%103213-32-75g,25g F100413Fmoc-O-叔丁基-L-谷氨酸 98%71989-18-95g,25g F100419Fmoc-L-谷氨酸 98%121343-82-65g,25g F100746N-Fmoc-N' -Boc-L-鸟氨酸 96%109425-55-01g,5g,25g F100759Fmoc-Val-OSu 97%130878-68-15g,25g F100801Fmoc-L-天冬氨酸 98%119062-05-41g,5g,25g,100g F100805Fmoc-L-缬氨酸 98%68858-20-85g,25g,100g F100808Fmoc-L-亮氨酸 98%35661-60-05g,25g,100g F101115FMOC-L-炔丙基甘氨酸 98%198561-07-81g,5g,250mg F101121FMOC-D-炔丙基甘氨酸 96%220497-98-31g,250mg F101195Fmoc-D-烯丙基甘氨酸 96%170642-28-11g,250mgF101202FMOC-D-3-(4-吡啶基)-丙氨酸 98%205528-30-91g,5g F101214Fmoc-3-(3-吡啶基)-L-丙氨酸 98%175453-07-31g,5g,250mg F101220FMOC-L-3-(2-吡啶基)-丙氨酸 97%185379-40-21g,250mg F101223FMOC-D-3-(2-吡啶基)-丙氨酸 98%185379-39-91g,5g F101459Fmoc-2-氨基异丁酸 97%94744-50-05g,25g F101574FMOC-L-4-甲基苯丙氨酸 98%199006-54-71g,250mg F101598FMOC-L-3-甲基苯丙氨酸 98%211637-74-01g,250mg F101600FMOC-D-3-甲基苯丙氨酸 98%352351-64-51gBoc修饰的氨基酸及氨基酸衍生物列表td style="padding-left: 12px "98%货号品名规格cas号包装 B100726BOC-O-苄基-L-酪氨酸 98%2130-96-35g,25g,100g B100799Boc-L-谷氨酰胺 98%13726-85-75g,25gB101207BOC-D-3-(3-吡啶基)-丙氨酸 98%98266-33-21g,5g,250mg B101451BOC-D-丙氨酸 98%7764-95-65g,25g B101478Boc-D-酪氨酸 70642-86-31g,5g,25g,100g B101548BOC-L-4-甲基苯丙氨酸 98%80102-26-71g,5g,250mg B101595BOC-L-3-甲基苯丙氨酸 98%114873-06-21g,5g B101597BOC-D-3-甲基苯丙氨酸 98%114873-14-21g,5g B101616BOC-L-2-甲基苯丙氨酸 98%114873-05-11g B101623BOC-D-2-甲基苯丙氨酸 98%80102-29-01g B101627BOC-D-4-溴苯丙氨酸 98%79561-82-31g B101633BOC-L-2-溴苯丙氨酸 98%261165-02-0500mg B101661BOC-L-3,4-二氯苯丙氨酸 98%80741-39-51g,5g,250mg B101686BOC-L-2-氯苯丙氨酸 98%114873-02-81g,5g B101696BOC-D-2-氯苯丙氨酸 98%80102-23-45g B102424Boc-L-脯氨酸酰胺 97%35150-07-31g,5g B102427N-BOC-L-苯丙氨醛 97%72155-45-41g,250mg B102428Boc-L-脯氨醛 97%69610-41-91g,5g B1024361-(Boc-氨基)环戊烷羧酸 98%35264-09-61g,5g B102447N(&alpha )-Boc-L-2,3-二氨丙酸 97%73259-81-11g,5g B102996BOC-L-异亮氨酸 99%13139-16-75g,25g,100g B103072N-Boc-N' -Cbz-L-赖氨酸 98%2389-45-95g,25g,100g B103084N-Boc-4-氧-L-脯氨酸甲酯 97%102195-80-21g,5g,250mg B103160(S)-N-BOC-4-溴苯丙氨酸 98%62129-39-91g,5g,25g更多产品请访问阿拉丁官网



















 我要推广仪器
我要推广仪器
 下载APP
下载APP