Edman降解法是测定蛋白质序列的经典方法,该方法由瑞士生物化学家佩尔维克托于1950年创立。Edman降解法通常是以周期的形式来表征。对于一个完整的周期,异硫氰酸苯酯标记上指定肽段的N末端,环化,之后被标记的氨基酸在酸性条件下从肽链中游离出来,进一步酸化后形成一个更加稳定的乙内酰苯硫脲-氨基酸(PTH-AA)衍生物。PTH-AA衍生物可以通过高效液相色谱鉴定。埃德曼降解周期就是通过反复地将多肽的氨基酸依次降解下来,从而鉴定氨基酸序列。 1982年,美国应用生物系统(ABI)公司了将第一台商用自动多肽测序仪推向市场,并被实践证明是可靠和耐用的。实际上,在过去几十年中,自动多肽测序仪只有屈指可数的一些改进,但此类自动肽测序仪仍是测定肽序列的黄金标准。然而,串联质谱的使用已经成为日益重要的肽序列测定工具。质谱,特别是串联飞行时间(TOF-TOF 和QTOF)质谱仪可以对少量样本进行更快速的分析。 与质谱相比,自动Edman降解测序的明显优势在于:已被化学验证的精确性、系统初始投资较小 然而,它的缺点是:分析时间较长、测得序列数有限,典型的测量长度范围是20~50个氨基酸序列。而对于低丰度的肽、无法获得N末端的肽,或者需要更短的分析时间,质谱则是理想的工具。但是,质谱价格高且无法区分同分异构的氨基酸。 虽然美国应用生物系统(ABI)公司主导了自动多肽测序仪的市场,但由于市场疲软,ABI于2008年6月停止生产自动多肽测序仪。然而岛津公司看准了机会,在2009年匹兹堡会议上将其PPSQ自动多肽测序仪推向北美市场,PPSQ已在日本广泛应用超过20年。另一方面,对用质谱测定蛋白质序列的方法改进的需求保持旺盛,布鲁克道尔顿公司最近推出了Edmass Micro MALDI-TOF和 Edmass Ultra TOF-TOF 系统,这是专门为没有质谱使用经验的多肽化学家们设计的。 自动多肽测序仪的市场主要来自于科研机构,但过去几年中自动多肽测序仪的市场需求增长处于某种停滞状态,尤其是质谱变得容易获得。但是,随着肽自动测序仪在连续使用过程中的老化,Edamn化学家们正在准备购买新设备。售后服务、附件、耗材及试剂有望在短期内对自动多肽测序仪市场产生推动作用。
各位大侠,给支个招,哪里可以既做高效液相又蛋白测序啊?我的样品现在是两个成分混在一起,分子量又都很小,我想测序,可最好是先用HPLC分开再来测,哪家公司或者研究所提供这方面的服务啊?
听说蛋白酶解肽段二级质谱图可以通过de novo方法测序,请问,除了手工解析,用哪些软件可以快速鉴定辅助解析
大家好,氨基酸分析仪与蛋白质测序仪有主要区别在什么地方呢?目前实验室需要进行氨基酸的测序分析,究竟买一台蛋白质测序仪好呢,还是氨基酸分析仪好呢?价格大概有多少呢?这些仪器有没有国产的呢 QQ:2392795357
蛋白质结构分析:制备、鉴定与微量测序
请问生物育种、基因测序、蛋白质 这些方面有哪些仪器啊?这些有关生物学方面的知识我一点都不明白。就连这个问题问的对不对,我都不知道!
大家好,我们这里现有一个分子量为11kd的蛋白质想要对其进行二级结构的核磁测序,但由于分子量相对比较大,我联系了国内的很多大学和科研机构都不能完成,因为我赶着毕业所以急需可以做这种检测的单位. 我希望大家可以向我提供对外可以做这么大分子量蛋白质核磁测序的单位,我们愿意付费对该蛋白质进行核磁测序. 对您的帮助表示万分感谢!! 谢谢谢谢谢谢!!!! 我的邮箱:lzjqq001@yahoo.com.cn
传统的蛋白鉴定方法,如免疫印迹法、内肽的化学测序、已知或未知蛋白的comigration分析,或者在一个有机体中有意义的基因的过表达通常耗时、耗力,不适合高流通量的筛选。 目前,所选用的技术包括对于蛋白鉴定的图象分析、微量测序、进一步对肽片段进行鉴定的氨基酸组分分析和与质谱相关的技术。1 图象分析技术(Image analysis)“满天星”式的2-DE图谱分析不能依靠本能的直觉,每一个图象上斑点的上调、下调及出现、消失,都可能在生理和病理状态下产生,必须依靠计算机为基础的数据处理,进行定量分析。 在一系列高质量的2-DE凝胶产生(低背景染色,高度的重复性)的前提下,图象分析包括斑点检测、背景消减、斑点配比和数据库构建。 首先,采集图象通常所用的系统是电荷耦合CCD(charge coupled device)照相机;激光密度仪(laser densitometers)和Phospho或Fluoroimagers,对图象进行数字化。 并成为以象素(pixels)为基础的空间和网格。 其次,在图象灰度水平上过滤和变形,进行图象加工,以进行斑点检测。 利用Laplacian,Gaussian,DOG(difference of Gaussians) opreator使有意义的区域与背景分离,精确限定斑点的强度、面积、周长和方向。图象分析检测的斑点须与肉眼观测的斑点一致。 在这一原则下,多数系统以控制斑点的重心或最高峰来分析,边缘检测的软件可精确描述斑点外观,并进行边缘检测和邻近分析,以增加精确度。 通过阈值分析、边缘检测、销蚀和扩大斑点检测的基本工具还可恢复共迁移的斑点边界。 以PC机为基础的软件Phoretix-2D正挑战古老的Unix为基础的2-D分析软件包。 第三,一旦2-DE图象上的斑点被检测,许多图象需要分析比较、增加、消减或均值化。 由于在2-DE中出现100%的重复性是很困难的,由此凝胶间的蛋白质的配比对于图象分析系统是一个挑战。 IPG技术的出现已使斑点配比变得容易。 因此,较大程度的相似性可通过斑点配比向量算法在长度和平行度观测。 用来配比的著名软件系统包括Quest,Lips,Hermes,Gemini等,计算机方法如相似性、聚类分析、等级分类和主要因素分析已被采用,而神经网络、子波变换和实用分析在未来可被采用。 配比通常由一个人操作,其手工设定大约50个突出的斑点作为“路标”,进行交叉配比。 之后,扩展至整个胶。例如:精确的PI和MW(分子量)的估计通过参考图上20个或更多的已知蛋白所组成的标准曲线来计算未知蛋白的PI和MW。 在凝胶图象分析系统依据已知蛋白质的pI值产生PI网络,使得凝胶上其它蛋白的PI按此分配。 所估计的精确度大大依赖于所建网格的结构及标本的类型。 已知的未被修饰的大蛋白应该作为标志,变性的修饰的蛋白的PI估计约在±0。25个单位。 同理,已知蛋白的理论分子量可以从数据库中计算,利用产生的表观分子量的网格来估计蛋白的分子量。 未被修饰的小蛋白的错误率大约30%,而翻译后蛋白的出入更大。 故需联合其他的技术完成鉴定。2 微量测序(microsequencing)蛋白质的微量测序已成为蛋白质分析和鉴定的基石,可以提供足够的信息。 尽管氨基酸组分分析和肽质指纹谱(PMF)可鉴定由2-DE分离的蛋白,但最普通的N-末端Edman降解仍然是进行鉴定的主要技术。 目前已实现蛋白质微量测序的自动化。 首先使经凝胶分离的蛋白质直接印迹在PVDF膜或玻璃纤维膜上,染色、切割,然后直接置于测序仪中,可用于subpicomole水平的蛋白质的鉴定。 但有几点需注意:Edman降解很缓慢,序列以每40 min 1个氨基酸的速率产生;与质谱相比,Edman降解消耗大;试剂昂贵,每个氨基酸花费
几种常用的蛋白鉴定方法传统的蛋白鉴定方法,如免疫印迹法、内肽的化学测序、已知或未知蛋白的comigration分析,或者在一个有机体中有意义的基因的过表达通常耗时、耗力,不适合高流通量的筛选。 目前,所选用的技术包括对于蛋白鉴定的图象分析、微量测序、进一步对肽片段进行鉴定的氨基酸组分分析和与质谱相关的技术。1 图象分析技术(Image analysis)“满天星”式的2-DE图谱分析不能依靠本能的直觉,每一个图象上斑点的上调、下调及出现、消失,都可能在生理和病理状态下产生,必须依靠计算机为基础的数据处理,进行定量分析。 在一系列高质量的2-DE凝胶产生(低背景染色,高度的重复性)的前提下,图象分析包括斑点检测、背景消减、斑点配比和数据库构建。 首先,采集图象通常所用的系统是电荷耦合CCD(charge coupled device)照相机;激光密度仪(laser densitometers)和Phospho或Fluoroimagers,对图象进行数字化。 并成为以象素(pixels)为基础的空间和网格。 其次,在图象灰度水平上过滤和变形,进行图象加工,以进行斑点检测。 利用Laplacian,Gaussian,DOG(difference of Gaussians) opreator使有意义的区域与背景分离,精确限定斑点的强度、面积、周长和方向。图象分析检测的斑点须与肉眼观测的斑点一致。 在这一原则下,多数系统以控制斑点的重心或最高峰来分析,边缘检测的软件可精确描述斑点外观,并进行边缘检测和邻近分析,以增加精确度。 通过阈值分析、边缘检测、销蚀和扩大斑点检测的基本工具还可恢复共迁移的斑点边界。 以PC机为基础的软件Phoretix-2D正挑战古老的Unix为基础的2-D分析软件包。 第三,一旦2-DE图象上的斑点被检测,许多图象需要分析比较、增加、消减或均值化。 由于在2-DE中出现100%的重复性是很困难的,由此凝胶间的蛋白质的配比对于图象分析系统是一个挑战。 IPG技术的出现已使斑点配比变得容易。 因此,较大程度的相似性可通过斑点配比向量算法在长度和平行度观测。 用来配比的著名软件系统包括Quest,Lips,Hermes,Gemini等,计算机方法如相似性、聚类分析、等级分类和主要因素分析已被采用,而神经网络、子波变换和实用分析在未来可被采用。 配比通常由一个人操作,其手工设定大约50个突出的斑点作为“路标”,进行交叉配比。 之后,扩展至整个胶。例如:精确的PI和MW(分子量)的估计通过参考图上20个或更多的已知蛋白所组成的标准曲线来计算未知蛋白的PI和MW。 在凝胶图象分析系统依据已知蛋白质的pI值产生PI网络,使得凝胶上其它蛋白的PI按此分配。 所估计的精确度大大依赖于所建网格的结构及标本的类型。 已知的未被修饰的大蛋白应该作为标志,变性的修饰的蛋白的PI估计约在±0。25个单位。 同理,已知蛋白的理论分子量可以从数据库中计算,利用产生的表观分子量的网格来估计蛋白的分子量。 未被修饰的小蛋白的错误率大约30%,而翻译后蛋白的出入更大。 故需联合其他的技术完成鉴定。2 微量测序(microsequencing)蛋白质的微量测序已成为蛋白质分析和鉴定的基石,可以提供足够的信息。 尽管氨基酸组分分析和肽质指纹谱(PMF)可鉴定由2-DE分离的蛋白,但最普通的N-末端Edman降解仍然是进行鉴定的主要技术。 目前已实现蛋白质微量测序的自动化。 首先使经凝胶分离的蛋白质直接印迹在PVDF膜或玻璃纤维膜上,染色、切割,然后直接置于测序仪中,可用于subpicomole水平的蛋白质的鉴定。 但有几点需注意:Edman降解很缓慢,序列以每40 min 1个氨基酸的速率产生;与质谱相比,Edman降解消耗大;试剂昂贵,每个氨基酸花费3~4$。 这都说明泛化的Edman降解蛋白质不适合分析成百上千的蛋白质。 然而,如果在一个凝胶上仅有几个有意义的蛋白质,或者如果其他技术无法测定而克隆其基因是必需的,则需要进行泛化的Edman降解测序。近来,应用自动化的Edman降解可产生短的N-末端序列标签,这是将质谱的序列标签概念用于Edman降解,业已成为一种强有力的蛋白质鉴定。 当对Edman的硬件进行简单改进,以迅速产生N-末端序列标签达10~20个/d,序列检签将适于在较小的蛋白质组中进行鉴定。若联合其他的蛋白质属性,如氨基酸组分分析、肽质质量、表现蛋白质分子量、等电点,可以更加可信地鉴定蛋白质。 选择BLAST程序,可与数据库相配比。 目前,采用一种Tagldent的检索程序,还可以进行种间比较鉴定,又提高了其在蛋白质组研究中的作用。3 与质谱(mass spectrometry)相关的技术质谱已成为连接蛋白质与基因的重要技术,开启了大规模自动化的蛋白质鉴定之门。 用来分析蛋白质或多肽的质谱有两个主要的部分,1)样品入机的离子源,2)测量被介入离子的分子量的装置。 首先是基质辅助激光解吸附电离飞行时间质谱(MALDI-TOF)为一脉冲式的离子化技术。 它从固相标本中产生离子,并在飞行管中测其分子量。 其次是电喷雾质谱(ESI-MS),是一连续离子化的方法,从液相中产生离子,联合四极质谱或在飞行时间检测器中测其分子量。 近年来,质谱的装置和技术有了长足的进展。在MALDI-TOF中,最重要的进步是离子反射器(ion reflectron)和延迟提取(delayed ion extraction),可达相当精确的分子量。 在ESI-MS中,纳米级电雾源(nano-electrospray source)的出现使得微升级的样品在30~40 min内分析成为可能。将反相液相色谱和串联质谱(tandem MS)联用,可在数十个picomole的水平检测;若利用毛细管色谱与串联质谱联用,则可在低picomole到高femtomole水平检测;当利用毛细管电泳与串联质谱连用时,可在小于femtomole的水平检测。 甚至可在attomole水平进行。 目前多为酶解、液相色谱分离、串联质谱及计算机算法的联合应用鉴定蛋白质。 下面以肽质指纹术和肽片段的测序来说明怎样通过质谱来鉴定蛋白质。(1)肽质指纹术(peptide mass fingerprint, PMF)由Henzel等人于1993年提出。 用酶(最常用的是胰酶)对由2-DE分离的蛋白在胶上或在膜上于精氨酸或赖氨酸的C-末端处进行断裂,断裂所产生的精确的分子量通过质谱来测量(MALDI-TOF-MS,或为ESI-MS),这一技术能够完成的肽质量可精确到0。1个分子量单位。 所有的肽质量最后与数据库中理论肽质量相配比(理论肽是由实验所用的酶来“断裂”蛋白所产生的)。 配比的结果是按照数据库中肽片段与未知蛋白共有的肽片段数目作一排行榜,“冠军”肽片段可能代表一个未知蛋白。若冠亚军之间的肽片段存在较大差异,且这个蛋白可与实验所示的肽片段覆盖良好,则说明正确鉴定的可能性较大。(2)肽片段(peptide fragment)的部分测序肽质指纹术对其自身而言,不能揭示所衍生的肽片段或蛋白质。 为进一步鉴定蛋白质,出现了一系列的质谱方法用来描述肽片段。 用酶或化学方法从N-或C-末端按顺序除去氨基酸,形成梯形肽片段(ladder peptide)。 首先以一种可控制的化学模式从N-末端降解,可产生大小不同的一系列的梯形肽片段,所得一定数目的肽质量由MALDI-TOF-MS测量。 另一种方法涉及羧基肽酶的应用,从C-末端除去不同数目的氨基酸形成肽片段。 化学法和酶法可产生相对较长的序列,其分子量精确至以区别赖氨酸(128。09)和谷氨酰胺(128。06)。 或者,在质谱仪内应用源后衰变(post-source decay, PSD)和碰撞诱导解离(collision-induced dissociation, CID),目的是产生包含有仅异于一个氨基酸残基质量的一系列肽峰的质谱。 因此,允许推断肽片段序列。 肽片段PSD的分析在MALDI反应器上能产生部分序列信息。 首先进行肽质指纹鉴定。 之后,一个有意义的肽片段在质谱仪被选作“母离子”,在飞行至离子反应器的过程中降解为“子离子”。 在反应器中,用逐渐降低的电压可测量至检测器的不同大小的片段。 但经常产生不完全的片段。 现在用肽片段来测序的方法始于70年代末的CID,可以一个三联四极质谱ESI-MS或MALDI-TOF-MS联合碰撞器内来完成。 在ESI-MS中,由电雾源产生的肽离子在质谱仪的第一个四极质谱中测量,有意义的肽片段被送至第二个四极质谱中,惰性气体轰击使其成为碎片,所得产物在第三个四极质谱中测量。 与MALDI-PSD相比,CID稳定、强健、普遍,肽离子片段基本沿着酰胺键的主架被轰击产生梯形序列。 连续的片段间差异决定此序列在那一点的氨基酸的质量。 由此,序列可被推测。 由CID图谱还可获得的几个序列的残基,叫做“肽序列标签”。 这样,联合肽片段母离子的分子量和肽片段距N- C端的距离将足以鉴定一个蛋白质。4 氨基酸组分分析1977年首次作为鉴定蛋白质的一种工具,是一种独特的“脚印”技术。 利用蛋白质异质性的氨基酸组分特征,成为一种独立于序列的属性,不同于肽质量或序列标签。 Latter首次表明氨基酸组分的数据能用于从2-DE凝胶上鉴定蛋白质。 通过放射标记的氨基酸来测定蛋白质的组分,或者将蛋白质印迹到PVDF膜上,在15
传统的蛋白鉴定方法,如免疫印迹法、内肽的化学测序、已知或未知蛋白的comigration分析,或者在一个有机体中有意义的基因的过表达通常耗时、耗力,不适合高流通量的筛选。 目前,所选用的技术包括对于蛋白鉴定的图象分析、微量测序、进一步对肽片段进行鉴定的氨基酸组分分析和与质谱相关的技术。1 图象分析技术(Image analysis)“满天星”式的2-DE图谱分析不能依靠本能的直觉,每一个图象上斑点的上调、下调及出现、消失,都可能在生理和病理状态下产生,必须依靠计算机为基础的数据处理,进行定量分析。 在一系列高质量的2-DE凝胶产生(低背景染色,高度的重复性)的前提下,图象分析包括斑点检测、背景消减、斑点配比和数据库构建。 首先,采集图象通常所用的系统是电荷耦合CCD(charge coupled device)照相机;激光密度仪(laser densitometers)和Phospho或Fluoroimagers,对图象进行数字化。 并成为以象素(pixels)为基础的空间和网格。 其次,在图象灰度水平上过滤和变形,进行图象加工,以进行斑点检测。 利用Laplacian,Gaussian,DOG(difference of Gaussians) opreator使有意义的区域与背景分离,精确限定斑点的强度、面积、周长和方向。图象分析检测的斑点须与肉眼观测的斑点一致。 在这一原则下,多数系统以控制斑点的重心或最高峰来分析,边缘检测的软件可精确描述斑点外观,并进行边缘检测和邻近分析,以增加精确度。 通过阈值分析、边缘检测、销蚀和扩大斑点检测的基本工具还可恢复共迁移的斑点边界。 以PC机为基础的软件Phoretix-2D正挑战古老的Unix为基础的2-D分析软件包。 第三,一旦2-DE图象上的斑点被检测,许多图象需要分析比较、增加、消减或均值化。 由于在2-DE中出现100%的重复性是很困难的,由此凝胶间的蛋白质的配比对于图象分析系统是一个挑战。 IPG技术的出现已使斑点配比变得容易。 因此,较大程度的相似性可通过斑点配比向量算法在长度和平行度观测。 用来配比的著名软件系统包括Quest,Lips,Hermes,Gemini等,计算机方法如相似性、聚类分析、等级分类和主要因素分析已被采用,而神经网络、子波变换和实用分析在未来可被采用。 配比通常由一个人操作,其手工设定大约50个突出的斑点作为“路标”,进行交叉配比。 之后,扩展至整个胶。例如:精确的PI和MW(分子量)的估计通过参考图上20个或更多的已知蛋白所组成的标准曲线来计算未知蛋白的PI和MW。 在凝胶图象分析系统依据已知蛋白质的pI值产生PI网络,使得凝胶上其它蛋白的PI按此分配。 所估计的精确度大大依赖于所建网格的结构及标本的类型。 已知的未被修饰的大蛋白应该作为标志,变性的修饰的蛋白的PI估计约在±0。25个单位。 同理,已知蛋白的理论分子量可以从数据库中计算,利用产生的表观分子量的网格来估计蛋白的分子量。 未被修饰的小蛋白的错误率大约30%,而翻译后蛋白的出入更大。 故需联合其他的技术完成鉴定。
在做氨基酸测定时,用异硫氰酸苯酯做衍生试剂,多余的衍生试剂要除去,现请教各位氨基酸与异硫氰酸苯酯反应方程式是什么样的,在衍生1umol/ml氨基酸单标时应用的100mmol/l异硫氰酸苯酯衍生试剂的量是多少?三乙胺在反应中起什么作用,在反应中应用的1mol/l的三乙胺的量是多少?另外在反应中用正己烷进行萃取,那么加入正己烷的量又是多少呢?
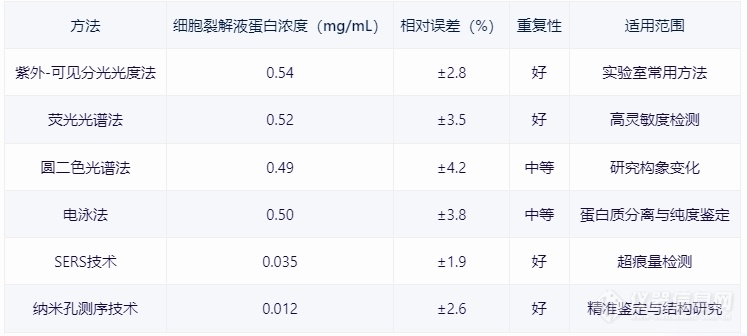
[b][font=宋体]一、引言[/font][/b][font=宋体] [/font][font=宋体]蛋白质浓度测定是生物化学实验中的一项基本操作,对于了解蛋白质的性质、评估实验效果以及开展生物学研究具有重要意义。本文将详细介绍蛋白浓度测定的方法、技术原理、最新进展及其在生物医学研究中的应用。[/font][font=宋体] [/font][b][font=宋体]二、蛋白浓度测定方法概述[/font][/b][font=宋体] [/font][font=宋体][font=宋体]①紫外[/font][font=Calibri]-[/font][font=宋体]可见分光光度法[/font][/font][font=宋体]原理:利用蛋白质中特定氨基酸(如酪氨酸、色氨酸)在紫外光区的吸收特性,通过测量吸光度值来计算蛋白质浓度。[/font][font=宋体] [/font][font=宋体]应用:适用于大多数蛋白质的测定,尤其在实验室中广泛使用。[/font][font=宋体] [/font][font=宋体]②荧光光谱法[/font][font=宋体]原理:利用荧光染料与蛋白质结合后产生的荧光光谱,通过测量荧光强度来计算蛋白质浓度。[/font][font=宋体] [/font][font=宋体]应用:适用于具有荧光特性的蛋白质,具有高灵敏度。[/font][font=宋体] [/font][font=宋体]③圆二色光谱法[/font][font=宋体]原理:利用圆二色光谱分析蛋白质的构象变化,从而推算蛋白质浓度。[/font][font=宋体] [/font][font=宋体]应用:常用于研究蛋白质的构象变化和稳定性。[/font][font=宋体] [/font][font=宋体]④电泳法[/font][font=宋体]原理:利用电泳技术将蛋白质分离,通过测量电泳带亮度或面积来计算蛋白质浓度。[/font][font=宋体] [/font][font=宋体]应用:常用于蛋白质分离和纯度鉴定。[/font][font=宋体] [/font][b][font=宋体]三、进展与技术优化[/font][/b][font=宋体] [/font][font=宋体][font=宋体]①表面增强拉曼散射([/font][font=Calibri]SERS[/font][font=宋体])技术[/font][/font][font=宋体]原理:利用金属表面增强拉曼散射效应,提高信号强度,从而提高检测灵敏度。[/font][font=宋体] [/font][font=宋体]应用:适用于痕量蛋白质的检测,具有高灵敏度和高分辨率。[/font][font=宋体] [/font][font=宋体]②纳米孔测序技术[/font][font=宋体]原理:利用纳米孔测序技术对蛋白质进行测序,通过电导变化检测蛋白质序列信息。[/font][font=宋体] [/font][font=宋体]应用:有助于蛋白质的精准鉴定和分子结构研究。[/font][font=宋体] [/font][font=宋体][b]四、[/b][/font][b][font=宋体]实际应用案例分析[/font][/b][font=宋体] [/font][align=center][img=蛋白浓度测定案例,690,310]https://ng1.17img.cn/bbsfiles/images/2024/01/202401231537106869_2656_5907840_3.png!w690x310.jpg[/img][font=宋体] [/font][/align][font=宋体][font=宋体]详情可以关注义翘神州更多[url=https://cn.sinobiological.com/resource/protein-review][b]蛋白资源[/b][/url]详情:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review[/font][/font][b][font=宋体][font=宋体]义翘神州:蛋白与抗体的专业引领者,欢迎通过百度搜索[/font][font=宋体]“义翘神州”与我们取得联系。[/font][/font][/b]
致病相关(PR)蛋白参与植物防御,其具有多种功能适应性,有助于抵抗各种病原体、提高环境胁迫耐受性。沙柳是一种生长迅速的柳树品种,可以耐受许多不利环境。中国林科院亚热带林业研究所卓仁英研究员课题组在[b][i]Environmental and Experimental Botany[/i][/b]上发表了一篇文章,题目为“Pathogenesis-related protein PR10 from [i]Salix matsudana [/i]Koidz exhibits resistance to salt stress in[i] [/i]transgenic [i]Arabidopsis thaliana[/i]”,主要探究PR蛋白在植物耐盐机制中起到的作用。前期的比较蛋白质组学分析表明:沙柳PR蛋白(SmPR10)较为丰富,经过100 mM NaCl处理后表达上调。本实验以沙柳为材料,克隆并鉴定了SmPR10基因,以验证其在耐盐性中的作用。SmPR10的氨基酸序列与紫苏柳和毛白杨的PR蛋白的序列同源性分别为98%和93%。SmPR10定位在拟南芥原生质体的胞质中,根的转录及蛋白水平较高,且100mM NaCl处理后表达上调。免疫定位分析发现,韧皮部纤维细胞和根木质部中特异性的检测到SmPR10。而且,SmPR10的异质过表达提高了转基因拟南芥的耐盐性,具体表现在根长度、根数量、Na[sup]+[/sup]流速、以及叶绿素含量、MDA含量、电导率等生理参数及SOD和POD酶活性水平。[align=center][img]https://mmbiz.qpic.cn/mmbiz_jpg/iaFShJzBuGDExem1Wsaria0uc8PElOtfBk8xxSu8n3s817ticGShAfXf1nbztqCgUODthZFzAqicXL736LrN3oSdww/640?wx_fmt=jpeg&wxfrom=5&wx_lazy=1[/img][/align][align=center][color=#888888]拟南芥根部Na[sup]+[/sup]流检测图[/color][/align][align=center][img]https://mmbiz.qpic.cn/mmbiz_png/iaFShJzBuGDExem1Wsaria0uc8PElOtfBkscqXvZYKA7ZZvOo8yMxAT3CnkhjazcdlWQJXoCupLI6hLTyppUHKBw/640?wx_fmt=png&wxfrom=5&wx_lazy=1[/img][/align][align=center][color=#888888][color=#888888]对照组、盐胁迫组拟南芥根部[/color]Na[sup]+[/sup][color=#888888]流结果。正值表示外排[/color][/color][/align]其中,Na[sup]+[/sup]流速利用基于非损伤微测技术(Non-invasive Micro-test Technology, NMT)的NMT活体生理检测仪 Physiolyzer[sup][/sup]进行检测,发现正常条件下,转基因组与野生对照组的Na[sup]+[/sup]外排相似。但是,100mM NaCl处理后,转基因组的Na[sup]+[/sup]外排明显增加。这说明:相比于野生型,SmPR10过表达幼苗根部的Na[sup]+[/sup]外排能力更高,从而更加耐盐。
求助:实验室除去异硫氰酸苯酯的方法有哪些?急!急!急!急!谢谢!
蛋白质是生物体中含量最高,功能最重要的生物大分子,存在于所有生物细胞,约占细胞干质量的50%以上, 作为生命的物质基础之一,蛋白质在催化生命体内各种反应进行、调节代谢、抵御外来物质入侵及控制遗传信息等方面都起着至关重要的作用,因此蛋白质也是生命科学中极为重要的研究对象。关于蛋白质的分析研究,一直是化学家及生物学家极为关注的问题,其研究的内容主要包括分子量测定,氨基酸鉴定,蛋白质序列分析及立体化学分析等。随着生命科学的发展,仪器分析手段的更新,尤其是质谱分析技术的不断成熟,使这一领域的研究发展迅速。 自约翰.芬恩(JohnB.Fenn)和田中耕一(Koichi.Tanaka)发明了对生物大分子进行确认和结构分析的方法及发明了对生物大分子的质谱分析法以来,随着生命科学及生物技术的迅速发展,生物质谱目前已成为有机质谱中最活跃、最富生命力的前沿研究领域之一[1]。它的发展强有力地推动了人类基因组计划及其后基因组计划的提前完成和有力实施。质谱法已成为研究生物大分子特别是蛋白质研究的主要支撑技术之一,在对蛋白质结构分析的研究中占据了重要地位[2]。 1.质谱分析的特点 质谱分析用于蛋白质等生物活性分子的研究具有如下优点:很高的灵敏度能为亚微克级试样提供信息,能最有效地与色谱联用,适用于复杂体系中痕量物质的鉴定或结构测定,同时具有准确性、易操作性、快速性及很好的普适性。 2.质谱分析的方法 近年来涌现出较成功地用于生物大分子质谱分析的软电离技术主要有下列几种:1)电喷雾电离质谱;2)基质辅助激光解吸电离质谱;3)快原子轰击质谱;4)离子喷雾电离质谱;5)大气压电离质谱。在这些软电离技术中,以前面三种近年来研究得最多,应用得也最广泛[3]。 3.蛋白质的质谱分析 蛋自质是一条或多条肽链以特殊方式组合的生物大分子,复杂结构主要包括以肽链为基础的肽链线型序列[称为一级结构]及由肽链卷曲折叠而形成三维[称为二级,三级或四级]结构。目前质谱主要测定蛋自质一级结构包括分子量、肽链氨基酸排序及多肽或二硫键数目和位置。 3.1蛋白质的质谱分析原理 以往质谱(MS)仅用于小分子挥发物质的分析,由于新的离子化技术的出现,如介质辅助的激光解析/离子化、电喷雾离子化,各种新的质谱技术开始用于生物大分子的分析。其原理是:通过电离源将蛋白质分子转化为[url=https://insevent.instrument.com.cn/t/Mp]气相[/url]离子,然后利用质谱分析仪的电场、磁场将具有特定质量与电荷比值(M/Z值)的蛋白质离子分离开来,经过离子检测器收集分离的离子,确定离子的M/Z值,分析鉴定未知蛋白质。 3.2蛋白质和肽的序列分析 现代研究结果发现越来越多的小肽同蛋白质一样具有生物功能,建立具有特殊、高效的生物功能肽的肽库是现在的研究热点之一。因此需要高效率、高灵敏度的肽和蛋白质序列测定方法支持这些研究的进行。现有的肽和蛋白质测序方法包括N末端序列测定的化学方法Edman法、C末端酶解方法、C末端化学降解法等,这些方法都存在一些缺陷。例如作为肽和蛋白质序列测定标准方法的N末端氨基酸苯异硫氰酸酯(phenylisothiocyanate)PITC分析法(即Edman法,又称PTH法),测序速度较慢(50个氨基酸残基/天);样品用量较大(nmol级或几十pmol级);对样品纯度要求很高;对于修饰氨基酸残基往往会错误识别,而对N末端保护的肽链则无法测序[4]。C末端化学降解测序法则由于无法找到PITC这样理想的化学探针,其发展仍面临着很大的困难。在这种背景下,质谱由于很高的灵敏度、准确性、易操作性、快速性及很好的普适性而倍受科学家的广泛注意。在质谱测序中,灵敏度及准确性随分子量增大有明显降低,所以肽的序列分析比蛋白容易许多,许多研究也都是以肽作为分析对象进行的。近年来随着电喷雾电离质谱(electrospray ionisation,ESI)及基质辅助激光解吸质谱(matrix assisted laser desorption/ionization,MALDI)等质谱软电离技术的发展与完善,极性肽分子的分析成为可能,检测限下降到fmol级别,可测定分子量范围则高达100000Da,目前基质辅助的激光解吸电离飞行时间质谱法(MALDI TOF MS)已成为测定生物大分子尤其是蛋白质、多肽分子量和一级结构的有效工具,也是当今生命科学领域中重大课题——蛋白质组研究所必不可缺的关键技术之一 [5] 。目前在欧洲分子生物实验室(EMBL)及美国、瑞士等国的一些高校已建立了MALDI TOF MS蛋白质一级结构(序列)谱库,能为解析FAST谱图提供极大的帮助,并为确证分析结果提供可靠的依据[6]。
我正在做细胞色素C 胰蛋白酶酶解后[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]测序,我想请教一下酶解的条件。酶的用量,时间,温度,pH,还有缓冲溶剂的配置
组蛋白乙酰化组蛋白修饰通过改变组蛋白与DNA的亲和性使染色质结构发生改变,进而影响转录因子与DNA序列的结合和基因表达,包括乙酰化、甲基化、磷酸化等,其中乙酰化是最重要的修饰方式之一,其主要发生在组蛋白H3赖氨酸(Lysine, Lys)的位点上,在癌症进展中发挥双重作用,既参与肿瘤抑制基因的沉默,又增强癌基因的表达[11],它受组蛋白乙酰转移酶(Histone acetyl transferase,HAT)和组蛋白去乙酰化酶(Histone deacetylase,HDAC)调控。HDAC可移去Lys残基上的乙酰基,增强组蛋白的正电性,DNA(本身带有负电荷)与组蛋白结合紧密,转录因子不易于DNA结合,抑制抑癌基因的转录,HAT作用则相反,二者动态平衡才能使组蛋白乙酰化维持在正常水平。表观遗传学改变通过调控基因转录平衡组蛋白乙酰化和去乙酰化,从而影响细胞周期、凋亡和分化相关蛋白的表达水平[12]。2.1 组蛋白乙酰化水平与SCLC发生发展密切相关 一项实验研究表明,乙酰化组蛋白H3在SCLC和NSCLC细胞中的表达有显著性差异,以前者表达较高。Notch信号通路是SCLC发生发展和化疗耐药的主要调节通路之一[13],具有肿瘤抑制作用。此研究中,Notch1在SCLC细胞系(除H69AR、SBC-3)中失活,其表达水平与组蛋白H3乙酰化有关。Notch1阳性表达的细胞系中乙酰化组蛋白H3富集在Notch1启动子区域,表达水平较高,Notch1阴性表达的细胞系中Notch1启动子周围的乙酰化组蛋白H3水平较低。这说明组蛋白去乙酰化是Notch1基因在SCLC中表观失活的原因[14]915-918。此外,组蛋白H3赖氨酸23(histone 3 lysine 23, H3K23)乙酰转移酶KAT6B在SCLC中失活,若其活性恢复可对SCLC产生抑制作用,它的乙酰化水平降低是SCLC发生的重要标志[15]。由此可见,组蛋白去乙酰化可以调控相关基因的表达从而促进SCLC发生发展。2.2 组蛋白去乙酰化酶抑制剂 HDAC在许多癌症中过表达,干扰其活性、抑制其功能是有效的治疗手段。组蛋白去乙酰化酶抑制剂(Histone deacetylase inhibitor, HDACI)是重要的表观调控药物,高效低毒,通过靶向阻断HDAC去乙酰化、促进组蛋白乙酰化发挥抗肿瘤作用。根据化学结构的不同,HDACIs分为异羟肟酸(异羟肟酸酯)、短链脂肪(脂肪族)酸、环状四肽、苯甲酰胺和Sirt抑制剂5类[16]。在单药和/或与传统化疗药物联合使用时,HDACI可阻滞细胞周期,抑制迁移和侵袭[17],诱导癌细胞分化、自噬[18]、凋亡,抗血管生成。当前,伏立诺他(Vorinostat ,SAHA)、罗米地辛(Romidepsin)、帕比司他(Panobinostat)等被批准用于血液系统恶性肿瘤的治疗[19]。丙戊酸(valproic acid ,VPA)作为HDACI可抑制SCLC细胞生长,诱导细胞凋亡,阻滞SCLC细胞周期于G1期。以上抑制作用是通过降低HDAC4表达,增加组蛋白H4乙酰化实现的。同时发现,VPA激活了SCLC中Notch1、Notch靶基因HES1和P21的Notch信号通路。此外,它还可以上调生长抑素受体II(somatostatinreceptor2,SSTR2)并增强受体靶向细胞毒素的抑制作用[20]。在经曲古抑菌素A (Trichostatin A ,TSA)处理后的SCLC细胞系中, Notch1启动子区域H3乙酰化水平增加,从而导致Notch1蛋白表达。此外,经TSA处理后,SCLC细胞黏附增加,上皮间质转化标志物表达减少,细胞增殖减少,细胞凋亡激活,可能与TSA诱导Notch1表达有关[14]916-918。这些研究成果为HDACI在 SCLC治疗中的应用提供了依据。为了达到最好治疗效果,药物用量、联合用药及使用顺序仍需深入研究。
[color=#444444]现在要测定一个植物花器官的分泌物成分,目前只知道里面有糖类和氨基酸,还有少量的蛋白和多肽,但是不清楚样品里面的蛋白具体是哪些物质或类型,只能估计有几丁质酶等一些大类,但也只是估计,是不是只能用[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]液质联用仪[/color][/url]测定?[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]液质联用[/color][/url]说是没有谱库,只能自己用标品去测,但是我又不清楚样品里具体物质是什么,不知道该用什么标品。不知道各位大神有没有别的方法?或者从哪里弄到谱库,或者质谱测序软件等能够鉴定出都是什么具体物质的方法,不用定量,能定性就行。在下感激不尽[/color]
含有一对或多对二硫键修饰的多肽:二硫键在蛋白质的结构稳定中起到重要作用,目前我们国肽生物已经能够为客户提供四对二硫键修饰的多肽。多对二硫键成环技术蛋白质和多肽类药物具有作用位点专一,疗效明确等优点,近年来,蛋白质和多肽类药物的研究和发展已经成为生物医药领域研究的一个热点。二硫键在维持多肽和蛋白质的空间立体结构及由此决定的生物活性中发挥着重要的作用。二硫键即为蛋白质或多肽分子中两个不同位点Cys的巯基(-SH)被氧化形成的S-S共价键。一条肽链上不同位置的氨基酸之间形成的二硫键,可以将肽链折叠成特定的空间结构。[align=center][img=,526,200]https://ng1.17img.cn/bbsfiles/images/2019/04/201904300953261874_7512_3531468_3.jpg!w526x200.jpg[/img][/align]多肽分子通常分子量较大,空间结构复杂,结构中形成二硫键时要求两个半胱氨酸在空间距离上接近。此外,多肽结构中还原态的巯基化学性质活泼,容易发生其他的副反应,而且肽链上其他侧链也可能会发生一系列修饰,因此,肽链进行修饰所选取的氧化剂和氧化条件是反应的关键因素,反应机理也比较复杂,既可能是自由基反应,也可能是离子反应。多肽的二硫键修饰中,分子内或者分子间一对二硫键的合成通常比较容易,反应条件有多种选择,比如空气氧化,DMSO氧化等温和的氧化过程,也可以采用H2O2,I2,汞盐等激烈的反应条件,反应产物也比较容易纯化分离,得到较高的纯度和产率。空气氧化法形成二硫键是多肽合成中最经典的方法,并且在早期的研究中取得了较好的结果。采用空气氧化法通常是将巯基处于还原态的多肽溶于水中,在近中性或弱碱性条件下(PH值6.5~10),反应24小时以上。为了降低分子之间二硫键形成的可能,该方法通常需要在低浓度条件下进行。碘氧化法在多肽合成中应用同样广泛,一般将多肽溶于25%的甲醇水溶液或30%的醋酸水溶液中,逐滴滴加10~15mol/L的碘进行氧化,反应15~40min。当肽链中含有对碘比较敏感的Tyr、Trp、Met和His的残基时,氧化条件要控制的更精确,氧化完后,立即加入维生素C或硫代硫酸钠除去过量的碘。当一条肽链上需要形成两对或两对以上的二硫键时,反应过程就变得相对复杂。在固相合成多肽之前,需要提前设计几对二硫键形成的顺序和方法路线,选择不同的侧链巯基保护基,利用其性质差异,分步氧化形成两对或多对二硫键。通常采用的巯基保护基有trt,Acm,Mmt,tBu,Bzl,Mob,Tmob等多种基团。我们分别列出两种以2-Cl树脂和Rink树脂为载体合成的多肽上多对二硫键形成路线:[align=center][img=,666,432]https://ng1.17img.cn/bbsfiles/images/2019/04/201904300953443844_9290_3531468_3.jpg!w666x432.jpg[/img][/align]二硫键的形成一直是多肽合成中的一个难点,经过不懈的研究和累积,我们公司已经具备相当成熟的多对二硫键成环技术,目前我们已经能够高成功率的合成三对和四对二硫键的多肽。不断克服实验困难,不停提高产品质量,不懈努力达到客户要求是我们国肽生物的不变宗旨。成功案例:固相合成序列DC*TSHNGAC*NHHSHC*C*SNVC*NTWAHLC*T,并对其成功进行3对二硫键修饰。HPLC分析:[align=center][img=,562,246]https://ng1.17img.cn/bbsfiles/images/2019/04/201904300954000045_2353_3531468_3.jpg!w562x246.jpg[/img][/align]MS分析:[align=center][img=,562,224]https://ng1.17img.cn/bbsfiles/images/2019/04/201904300954145791_912_3531468_3.jpg!w562x224.jpg[/img][/align][align=left]我们主要提供:多肽合成、定制多肽、同位素标记肽、人工胰岛素、磷酸肽、生物素标记肽、荧光标记肽(Cy3、Cy5、Fitc、AMC等)、目录肽、偶联蛋白(KLH、BSA、OVA等)、化妆品肽、多肽文库构建、抗体服务、糖肽、订书肽、药物肽、RGD环肽等。合肥国肽生物官网:http://www.bankpeptide.com欢迎咨询服务热线:17718122172;17718122684;17730030476;17718122397[/align]
现在,在实验研究基础上,借助多方面的生物信息学方法,可以快速高通量的预测和进行蛋白质鉴定蛋白翻译后修饰。分泌蛋白和膜相关蛋白附着于细胞膜上的或将被排泄出去的蛋白质是由细胞内质网膜上附着的核糖体合成。附着有核糖体的内质网被称为糙面型内质网。这类蛋白质都含有一个N-末端(或氨基端),我们称之为信号序列或信号肽。这个信号肽通常情况下含有13-36个主要疏水性残基,同时它含有多蛋白复合物,我们称之为信号识别粒子(SRP)。这种信号肽在通过内质网膜之后会被去除。信号肽的去除过程是在信号肽酶催化作用下完成的。含有一个信号肽的蛋白质被称为前蛋白,有别于原蛋白。然而,某些用于分泌的蛋白在分泌之后会进一步被蛋白水解,因此包含有原蛋白的序列。这类蛋白质被称为前原蛋白。蛋白水解性裂解许多蛋白质在翻译之后会经历水解性裂解过程。其中最为简单的形式是去除起始蛋氨酸。许多蛋白质合成了不活跃的前体细胞,这些细胞只能在合适的生理条件下通过限制性蛋白水解过程产生活性。在凝血过程中使用到的胰腺酶和酶类就是后者的例证。多肽去除时产生活性的不活跃的前体蛋白,我们称之为原蛋白。前原蛋白的翻译后加工过程的一个复杂的例子就是脑垂体分泌合成的前阿黑皮素原的裂解过程(有关前阿黑皮素原的讨论,见肽类激素页)。这类前原蛋白经过复杂的裂解,根据合成的前阿黑皮素原的细胞定位而不同,其路径也有所不同。另一个前原蛋白的例子就是胰岛素。由于胰岛素是由胰腺分泌的,因此它有一个前肽。随着含24个氨基酸的信号肽的裂解,这类蛋白也折叠成了胰岛素原。胰岛素原进一步分裂,产生活跃的胰岛素,它包含两个肽链,由二硫键进行连接。但仍有其他的蛋白(酶类)被合成为非活跃的前体细胞,被称为酶原。酶原在蛋白水解性裂解时会产生活性,在凝血串联蛋白质链的若干蛋白质中都会发生这种现象。甲基化作用蛋白翻译后的甲基化过程主要发生在氮原子和氧原子上。活性甲基供体是活性腺苷甲硫胺酸(SAM)。最常见的甲基化作用发生在赖氨酸残基的ε-amine上。脱氧核糖核酸组蛋白中赖氨酸残基的甲基化作用可调节核染色质结构,因此可调节其转录活性。赖氨酸原本被认为是一种常设共价标记,可提供长期信号,甚至包括转录记忆时的组蛋白依赖机制。然而,最近的临床研究表明赖氨酸甲基化作用与其他共价修饰体相似,作用时间短,并能通过反脱甲基化活动进行动态调节。最近的组学研究发现表明,赖氨酸残基的甲基化作用不仅发生在核染色质层面,而且还通过修订转录因子影响基因表达。组氨酸的咪唑环,精氨酸的胍基部分以及谷氨酸盐和天冬氨酸盐的R组酰胺(R-group amides )上,都发现了额外的氮甲基化作用。谷氨酸盐和天冬氨酸盐的R组羧化物也会发生氧甲基化作用并形成甲基酯。蛋白可能在半胱氨酸的R[
RIP技术(RNA Binding Protein Immunoprecipitation,RNA结合蛋白免疫沉淀),是研究细胞内RNA与蛋白结合情况的技术,是了解转录后调控网络动态过程的有力工具,能帮助我们发现miRNA的调节靶点。RIP这种新兴的技术运用针对目标蛋白的抗体把相应的RNA-蛋白复合物沉淀下来,然后经过分离纯化就可以对结合在复合物上的RNA进行分析。RIP可以看成是普遍使用的染色质免疫沉淀ChIP技术的类似应用,但由于研究对象是RNA-蛋白复合物而不是DNA-蛋白复合物,RIP实验的优化条件与ChIP实验不太相同(如复合物不需要固定,RIP反应体系中的试剂和抗体绝对不能含有RNA酶,抗体需经RIP实验验证等等)。RIP技术下游结合microarray技术被称为RIP-Chip,帮助我们更高通量地了解癌症以及其它疾病整体水平的RNA变化。Millipore基于磁珠的RIP实验流程RIP 实验基本原理:1. 用抗体或表位标记物捕获细胞核内或细胞质中内源性的RNA结合蛋白。2. 防止非特异性的RNA的结合。3. 免疫沉淀把RNA结合蛋白及其结合的RNA一起分离出来。4.结合的RNA序列通过microarray(RIP-Chip),定量RT-PCR或高通量测序(RIP-Seq)方法来鉴定。延伸阅读:95%的人类基因组并不编码基因,而是产生大量的非编码RNA,真正编码蛋白质的基因只占人类总基因组的约2%。这些非编码RNA在生命的生长发育的各个阶段都发挥着重要的调节作用,与艾滋病、白血病、糖尿病、畸形等多种病变密切相关,并且参与着干细胞和表观遗传学调控。而RNA-蛋白复合物驱动了几乎所有细胞过程的基因表达的转录后调控,包括剪接(splicing)、出核转运(nuclear export)、mRNA 稳定性以及蛋白转译过程,因此,对基因调控的了解就有赖于确定这些过程中RNA的结合的变化。因此,RNA研究也被越来越多的科学家重视起来,目前已经成为生命科学研究中一个炙手可热的领域,而RIP技术也逐渐成为RNA研究领域的一项常规方法,帮助我们了解越来越受关注的转录后调控网络。
最近在用异硫氰酸苯酯(PITC)检测氨基酸的时候出现了一个怪问题:17个氨基酸只有前面9种氨基酸可以出峰,剩下的氨基酸都不能出峰。液相条件什么都是一样的,换了新柱子问题仍然存在。一开始怀疑由于现在天气凉了衍生不充分,就放在25℃水浴中衍生2小时,问题仍然存在。让我很是苦恼。调整梯度程序、温度都不管用。液相也很正常。下面是我的液相条件:请各位高手帮忙啊!5色谱条件色谱柱: Sepax AA专用柱,4.6*250mm,由苏州赛分科技有限公司提供。检测波长:254nm。柱温:40℃。流动相A: 醋酸钠溶液:乙腈=93:7 (V/V)。流动相B: 乙腈:水=4:1(V/V)。进样量:5ul流速:1mL/min(注:分析结束后使用20%乙腈水溶液清洗,再用100%乙腈清洗后储存) 表1 梯度洗脱程序 时间/min 流动相A/% 流动相B/% 0 100 0 2 100 0 14 93 7 29 70 30 32 50 50 33 0 100 39 0 100 39.1 100 0 45 100 0 6 氨基酸标准溶液和样品溶液的衍生(设2个重复)6.1 17AA衍生准确量取氨基酸标准溶液400μl,置5.0ml塑料离心管中,加入三乙胺乙腈溶液(2.10)200μl,异硫氰酸苯酯乙腈溶液(2.11)200μl,混匀,室温放置1小时,然后加入正己烷800μl,振摇后放置10分钟,取下层溶液(PTC-AA),用一次性注射器吸取上清液用0.45μm针式过滤器过滤。准确量取样品溶液400μl,置5.0ml塑料离心管中,加入三乙胺乙腈溶液(2.10)200μl,异硫氰酸苯酯乙腈溶液(2.11)200μl,混匀,室温放置1小时,然后加入正己烷800μl,振摇后放置10分钟,取下层溶液(PTC-AA),用一次性注
什么是粗蛋白, 粗蛋白跟蛋白质又有什么区别,如何测量饲料中粗蛋白的含量,粗蛋白的含量高是不是一定代表着蛋白质的含量高。我想,当你看到这个题目时,肯定会联想到这一连串的问题中的其中几个。那么接下来,我就来详细介绍下粗蛋白的概念、粗蛋白测量和其他关于饲料中粗蛋白含量的问题。粗蛋白概念:粗蛋白不仅包括蛋白质这一物质,它涵盖的范围更广,包括含氮的全部物质。粗蛋白含量:下面我介绍几种常见物质的粗蛋白含量,仅供大家参考。薏苡仁 粗蛋白含量:13%-14%棉粕 粗蛋白含量:可达40%以上农大白早糯玉米 粗蛋白含量:3.41%蠡玉168 粗蛋白含量:9.63%台湾大青枣 粗蛋白含量:0.86%上文介绍了几种农产品或水果的粗蛋白含量情况,如果需要更多的资料,大家可自己查阅。粗蛋白测定:方法一:最简便也是最快键的方法,就是用蛋白质测定仪(参见www.top17.net/product/993.html)来测量。本标准参照采用ISO 5983—1979 《动物饲料──氮含量的测定和粗蛋白含量计算》。 1 主题内容与适用范围 本标准规定了饲料中粗蛋白含量的测定方法。 本标准适用于配合饲料、浓缩饲料和单一饲料。2 引用标准 GB 601 化学试剂 滴定分析(容量分析)用标准溶液的制备3 原理 凯氏法测定试样中的含氮量,即在催化剂作用下,用硫酸破坏有机物,使含氮物转化成硫酸铵。加入强碱进行蒸馏使氨逸出,用硼酸吸收后,再用酸滴定,测出氮含量,将结果乘以换算系数6.25,计算出粗蛋白含量。4 试剂4.1 硫酸(GB 625):化学纯,含量为98%,无氮。4.2 混合催化剂:0.4g硫酸铜,5个结晶水(GB 665),6g硫酸钾(HG 3—920)或硫酸钠(HG 3—908),均为化学纯,磨碎混匀。4.3 氢氧化钠(GB 629):化学纯,40%水溶液(m/V)。4.4 硼酸(GB 628):化学纯,2%水溶液(m/V)。4.5 混合指示剂:甲基红(HG 3—958)0.1%乙醇溶液,溴甲酚绿(HG 3—1220)0.5%乙醇溶液,两溶液等体积混合,在阴凉处保存期为三个月。4.6 盐酸标准溶液:邻苯二甲酸氢钾法标定,按GB 601制备。4.6.1 盐酸标准溶液:c(HCl)=0.1mol/L。8.3mL盐酸(GB 622,分析纯),注入 1 000mL蒸馏水中。4.6.2 盐酸标准溶液:c(HCl)=0.02mol/L。1.67mL盐酸(GB 622,分析纯),注入1 000mL蒸馏水中。4.7 蔗糖(HG 3—1001):分析纯。4.8 硫酸铵(GB 1396):分析纯,干燥。4.9 硼酸吸收液:1%硼酸水溶液1 000mL,加入0.1%溴甲酚绿乙醇溶液10mL,0.1%甲基红乙醇溶液7mL,4%氢氧化钠水溶液0.5mL,混合,置阴凉处保存期为一个月(全自动程序用)。5 仪器设备5.1 实验室用样品粉碎机或研钵。5.2 分样筛:孔径0.45mm(40目)。5.3 分析天平:感量0.0001g。5.4 消煮炉或电炉。5.5 滴定管:酸式,10、25mL。5.6 凯氏烧瓶:250mL。5.7 凯氏蒸馏装置:常量直接蒸馏式或半微量水蒸气蒸馏式。5.8 锥形瓶:150、250mL。5.9 容量瓶:100mL。5.10 消煮管:250mL。5.11 定氮仪:以凯氏原理制造的各类型半自动,全自动蛋白质测定仪。6 试样的选取和制备 选取具有代表性的试样用四分法缩减至200g,粉碎后全部通过40目筛,装于密封容器中,防止试样成分的变化。7 分析步骤7.1 仲裁法7.1.1 试样的消煮 称取试样0.5~1g(含氮量5~80mg)准确至0.0002g,放入凯氏烧瓶(5.6)中,加入6.4g混合催化剂(4.2),与试样混合均匀,再加入12mL硫酸(4.1)和2粒玻璃珠,将 凯氏烧瓶(5.6)置于电炉(5.4)上加热,开始小火,待样品焦化,泡沫消失后,再加强火力 (360~410℃)直至呈透明的蓝绿色,然后再继续加热,至少2h。7.1.2 氨的蒸馏(蒸馏步骤的检验见附录A)7.1.2.1 常量蒸馏法 将试样消煮液(7.1.1)冷却,加入60~100mL蒸馏水,摇匀,冷却。将蒸馏装置(5.7)的冷凝管末端浸入装有25mL硼酸(4.4)吸收液和2滴混合指示剂(4.5)的锥形瓶内。然后小心地向凯氏烧瓶(5.6)中加入50mL氢氧化钠溶液(4.3),轻轻摇动凯氏烧瓶(5.6),使溶液混匀后再加热蒸馏,直至流出液体积为100mL。降下锥形瓶,使冷凝管末端离开液面,继续蒸馏1~2min,并用蒸馏水冲洗冷凝管末端,洗液均需流入锥形瓶内,然后停止蒸馏。7.1.2.2 半微量蒸馏法 将试样消煮液(7.1.1)冷却,加入20mL蒸馏水,转入100mL容量瓶中,冷却后用水稀释至刻度,摇匀,做为试样分解液。将半微量蒸馏装置(5.7)的冷凝管末端浸入装有20mL硼酸(4.4)吸收液和2滴混合指示剂(4.5)的锥形瓶(5.8)内。蒸汽发生器 (5.7)的水中应加入甲基红指示剂数滴,硫酸数滴,在蒸馏过程中保持此液为橙红色,否则需补加硫酸。准确移取试样分解液10~20mL注入蒸馏装置(5.7)的反应室中,用少量蒸馏水冲洗进样入口,塞好入口玻璃塞,再加10mL氢氧化钠溶液(4.3),小心提起玻璃塞使之流入反应室,将玻璃塞塞好,且在入口处加水密封,防止漏气。蒸馏4min降下锥形瓶(5.8)使冷凝管末端离开吸收液面,再蒸馏1min,用蒸馏水冲洗冷凝管末端,洗液均流入锥形瓶内,然后停止蒸馏。 注:7.1.2.1和7.1.2.2蒸馏法测定结果相近,可任选一种。7.1.2.3 蒸馏步骤的检验 精确称取0.2g硫酸铵(4.8),代替试样,按7.1.2或7.2.2步骤进行操作,测得硫酸铵含氮量为21.19±0.2%,否则应检查加碱、蒸馏和滴定各步骤是否正确。 7.1.3 滴定 用7.1.2.1或7.1.2.2法蒸馏后的吸收液立即用0.1mol/L(4.6.1)或0.02mol/L(4.6.2)盐酸标准溶液滴定,溶液由蓝绿色变成灰红色为终点。7.2 推荐法7.2.1 试样的消煮 称取0.5~1g试样(含氮量5~80mg)准确至0.0002g,放入消化管中,加2片消化片(仪器自备)或6.4g混合催化剂(4.2),12mL硫酸(4.1),于420 ℃下在消煮炉上消化1h。取出放凉后加入30mL蒸馏水。7.2.2 氨的蒸馏 采用全自动定氮仪(5.11)时,按仪器本身常量程序进行测定。 采用半自动定氮仪(5.11)时,将带消化液的管子插在蒸馏装置上,以25mL硼酸(4.4)为吸收液,加入2滴混合指示剂(4.5),蒸馏装置(5.7)的冷凝管末端要浸入装有吸收液的锥形瓶内,然后向消煮管中加入50mL氢氧化钠溶液(4.3)进行蒸馏。蒸馏时间以吸收液体积达到100mL时为宜。降下锥形瓶,用蒸馏水冲洗冷凝管末端,洗液均需流入锥形瓶内。7.2.3 滴定 用0.1mol/L的标准盐酸溶液(4.6.1)滴定吸收液,溶液由蓝绿色变成灰红色为终点。8 空白测定 称取蔗糖0.5g,代替试样,按第7章进行空白测定,消耗0.1mol/L盐酸标准溶液(4.6.1)的体积不得超过0.2mL。消耗0.02mol/L盐酸标准溶液(4.6.2)体积不得超过0.3mL。9 分析结果的表述9.1 计算见下式:粗蛋白质(%)=(V2-V1)• c×0.0140×6.25/(m×V'/V) ×100 式中:V2── 滴定试样时所需标准酸溶液体积,mL; V1── 滴定空白时所需标准酸溶液体积,mL; c── 盐酸标准溶液浓度,mol/L; m── 试样质量,g; V── 试样分解液总体积,mL; V── 试样分解液蒸馏用体积,mL; 0.0140── 与1.00mL盐酸标准溶液〔c(HCl)=1.000mol/L〕相当的、以克表示的氮的质量。 6.25── 氮换算成蛋白质的平均系数。 9.2 重复性 每个试样取两个平行样进行测定,以其算术平均值为结果。 当粗蛋白质含量在25%以上时,允许相对偏差为1%。 当粗蛋白含量在10%~25%之间时,允许相对偏差为2%。 当粗蛋白质含量在10%以下时,允许相对偏差为3%。
http://img.dxycdn.com/trademd/upload/asset/meeting/2013/09/06/A1378379551.jpg 氨基酸是蛋白质的基础组成单位,通过研究蛋白质中氨基酸的性质和组成来预测蛋白质的结构和功能,蛋白质氨基酸残基组成分析主要是通过氨基酸分析仪来完成的,本文推荐了2个基于氨基酸组成进行蛋白质预测软件。基于氨基酸组成的蛋白质预测软件根据组成蛋白质的20种氨基酸的物理和化学性质可以辨析电泳等实验中的未知蛋白质,也可以分析已知蛋白质的物化性质。ExPASy工具包包涵的程序:AACompIdent:与把氨基酸序列在SWISS-PROT库中搜索不同,AACompIdent工具利用未知蛋白的氨基酸组成去确认具有相同组成的已知蛋白。该程序分析时需提交的相关信息包括:蛋白质的氨基酸组成、等电点pI和分子量(如果知道)、正确的物种分类及特别的关键词。此外,用户还需在六种氨基酸“组合”中作出选择,这影响到分析如何进行。例如,某种“组合”会把残基Asp/Asn(D/N)和Gln/Glu(Q/E)组合成 Asx(B)和Glx(Z);或者某种残基会在分析中被完全除去。对数据库中的每一个蛋白序列,算法会对其氨基酸组成与所查询的氨基酸组成的差异打分。由电子邮件返回的结果被组织成三级列表:第一张列表中的蛋白都基于特定的物种分类而不考虑pI和分子量;第二张列表包含了不考虑物种分类、pI和分子量的全体蛋白;第三张列表中的蛋白不但基于特定物种分类,并且将 pI和分子量也考虑在内。虽然计算所得结果各不相同,但零分表明了该序列与提出的组成完全相符。AACompSim:AACompIdent的一个变种,AACompSim提供类似的分析,但与前者以实验所得的氨基酸组成为依据进行搜索不同,后者使用SWISS-PROT中的序列为依据。有报道称,氨基酸组成在物种之间是十分保守的(Cordwell等,1995),并且通过分析氨基酸的组成,研究者能从低于25%序列相似性的蛋白之间发现弱相似性(Hobohm和Sander,1995)。因此,在“传统的”数据库搜索基础上辅以组成分析,能为蛋白质之间关系提供更多见解。PROSEARCH:PROPSEARCH也提供基于氨基酸组成的蛋白质辨识功能。用144种不同的物化性质来分析蛋白质,包括分子量、巨大残基的含量、平均疏水性、平均电荷等,把查询序列的这些属性构成的“查询向量”与SWISS-PROT和PIR中预先计算好的各个已知蛋白质的属性向量进行比较。这个工具能有效的发现同一蛋白质家族的成员。可以通过Web使用这个工具,用户只需输入查询序列本身。分子量搜索(MOWSE)分子量搜索(MolecularWeightSearch,MOWSE)算法利用了通过质谱(MS)技术获得的信息。利用完整蛋白质的分子量及其被特定蛋白酶消化后产物的分子量,一种未知蛋白质能被准确无误地确认,给出由若干实验才能决定的结果。由于未知蛋白无需再全部或部分测序,这一方法显著地减少了实验时间。MOWSE的输入是一个纯文本文件,包含一张实验测定的肽段列表,分子量范围在0.7到4.0Kda之间。计算过程基于在OWL非冗余蛋白质序列库中包含的信息。打分基于在一定分子量范围内蛋白中一个片段分子量出现的次数。输出的结果是得分最佳的30个蛋白的列表,包括它们在OWL中的条目名称、相符肽段序列、和其它统计信息。模拟研究得出在使用5个或更少输入肽段分子量时,准确率为99%。该搜索服务可通过向mowse@daresburg.ac.uk发送电子邮件实现。为获得更多关于查询格式的细节信息,可以相该地址发送电子邮件,并在消息正文中写上“help”这个词。蛋白质氨基酸组成分析用盐酸在110 ℃将蛋白或多肽水解成游离的氨基酸,用氨基酸分析仪测定各氨基酸的含量。采用经典的阳离子交换色谱分离、茚三酮柱后衍生法,对蛋白质水解液及各种游离氨基酸的组分含量进行分析。仪器基本结构同普通HPLC相似,但针对氨基酸分析进行了细节优化(例如氮气保护、惰性管路、在线脱气、洗脱梯度及柱温梯度控制等等)通常细分为两种系统:蛋白水解分析系统(钠盐系统)和游离氨基酸分析系统(锂盐系统),利用不同浓度和pH值的柠檬酸钠或柠檬酸锂进行梯度洗脱。其中钠盐系统一次最多分析约25种氨基酸,速度较快,基线平直度好;锂盐系统一次最多分析约50种氨基酸,速度较慢,基线一般不如钠盐系统好。分析效果:从目前已知的氨基酸分析方法比较来看,除灵敏度(即最低检测限)比HPLC柱前衍生方法稍低以外(HPLC:0.5 pmol;氨基酸分析仪:10 pmol),其他如分离度、重现性、操作简便性、运行成本等方面,都优于其他分析方法。蛋白质氨基酸残基组成分析的主要步骤包括:首先是蛋白被水解为氨基酸,其次是采用离子色谱等方法进行游离的氨基酸含量和组成的分析。总之利用蛋白可以分析氨基酸,利用氨基酸也可以研究蛋白质。
本文引用自cheney《蛋白胨和胰蛋白胨简介》蛋白胨是将肉、酪素或明胶用酸或蛋白酶水解后干燥而成的外观呈淡黄色的粉剂,具有肉香的特殊气息。蛋白质经酸、碱或蛋白酶分解后也可形成蛋白胨。蛋白胨富含有机氮化合物,也含有一些维生素和糖类。它可以作为微生物培养基的主要原料,在抗生素、医药工业、发酵工业、生化制品及微生物学科研等领域中的用量均很大。不同的生物体需要特定的氨基酸和多肽,因此存在着各种蛋白胨,一般来说,用于蛋白胨生产的蛋白包括动物蛋白(酪蛋白、肉类)和植物蛋白(豆类)等两种。能为微生物提供C源、N源、生长因子等营养物质。因此,蛋白胨从来源上可分为动物性蛋白胨和植物性蛋白胨。胰胨、肉胨、骨胨等都是动物性蛋白胨,而大豆蛋白胨等则是植物性蛋白胨。动物性来源的蛋白胨还有:蚕蛹蛋白胨、血液蛋白胨等。 不同来源的蛋白质和不同的水解条件,其水解物中组成可千差万别。所以胨往往是一个复杂的多肽混合物。可溶于水,过热不凝固,在饱和硫酸铵中不发生沉淀但可为蛋白质沉淀剂所沉淀。可用作微生物和动物细胞培养基、特种功能性食品和化妆品的配料,也有用作啤酒等产品的稳定剂。胰蛋白胨,又称胰酪蛋白胨(Casein Tryptone)、胰酶消化酪蛋白胨(Pancreatic digest of casein),是一种优质蛋白胨,是以新鲜牛肉和牛骨经胰酶消化,浓缩干燥而成的浅黄色粉末。具有色浅、易溶、透明、无沉淀等良好的物理性状。含有丰富的氮源、氨基酸等,可配制各种微生物培养基,用于细菌的培养、分离、增殖、鉴定,以及无菌试验培养基、厌氧菌培养基等细菌生化特性试验用培养基的配置。胰蛋白胨还广泛应用于高品质的抗生素、维生素、医药工业,氨基酸、有机酸、酶制剂、黄原胶等发酵工业,生化制品及微生物学科研等领域中的用量均很大,临床用于抗炎消肿,工业上用于皮革制造,生丝处理,食品加工。在国际市场上,胰蛋白胨也属于货紧价昂的短线品种之一。 胰酪蛋白胨质量标准及其检验标准: 常规各项理化指标: 1. 澄清度(磷酸盐、碱性沉淀):无沉淀、澄清 2. 2%水溶液:透明 3. 酸碱度:6-7 4. 氨基氮:≥3% 5. 色氨酸:≥0.8% 6. 胨含量:≥80% 7. 总氮:≥13% 8. 水份:≤5% 9. 灰份:≤6% 10. 氯化钠:≤0.2%胰蛋白胨特指用胰蛋白酶酶解酪蛋白生成的蛋白胨产物,与一般蛋白胨的区别在于酶解工艺处理上,属于水解度更高、胨分子量更小更均衡的蛋白胨。
[摘要] 质谱对于现代蛋白质化学研究是一项重要的技术。也可用于蛋白组分析,在同一时间监控上千种蛋白的表达情况。首先蛋白质混合物可以用双向凝胶电泳分开,再用质谱及随后的蛋白质数据库检索对单个蛋白进行鉴定和分析。最近几年,质谱领域取得了令人鼓舞的进展,使得全自动、高通量的蛋白检测成为可能。质谱也可以用来分析蛋白的转录后调节以及研究蛋白质复合体。本文将介绍目前蛋白组研究中质谱方法的应用并对其优缺点进行讨论。关键词: 质谱;二维电泳(2-DE);蛋白组分析MALDI-TOF1 前 言蛋白组学是研究生物特定组织或细胞内表达的所有蛋白质成分,蛋白质组的一门学科。蛋白组分析一般是用双向凝胶电泳(2-DE),质谱(MS)以及数据检索来完成。2-DE要回顾到20世纪70年代初[1],但是用质谱技术对蛋白质进行鉴定和分析是在近十年内才成为可能。其中最重要的一个原因是新的软离子技术即基质辅助激光解析电离(MALDI)[2]和电喷雾电离(ESI)[3,4]技术的发展和应用,以及样本制备技术和质谱仪的发展等。从数据库中获得成指数上升的序列信息也促进了质谱技术的发展。跟转录组学中的全自动DNA微阵列技术相比,蛋白组分析需要更多的手工操作,尤其在2-DE上会出现很多问题[5,6]。尽管存在很多困难,蛋白组学的重要性还是公认的。DNA微阵列技术是建立在对mRNA稳定水平的检测上,然而蛋白组学研究的对象是细胞中最活跃的因子——蛋白质。最近有研究证明,mRNA的表达和蛋白的水平并非具有相关性[7,8]。蛋白质有转录后修饰过程,并会以不同的形式出现。而这些修饰过程是相应的DNA测序技术所不能预测的。质谱技术在蛋白质修饰分析过程中具有重要的作用。2 质谱对蛋白质的检测质谱仪是由离子源,质量分析,离子检测器,以及数据检索等单元组成。首先,被分析的分子在离子源中被离子化,然后在质量分析器里根据不同的质荷比分开,再对分开的离子进行检测。随着MALDI和EIS的出现,质谱技术已经广泛用来分析蛋白质。质量分析器有很多种,最常用的方法是将飞行时间检测器(TOF)与MALDI或三级四级串联质谱仪联用,或者将四级杆-TOF,离子阱等连接到ESI上。蛋白组分析中,可以用两种不同的方法检测电泳分离后的单个蛋白质。最简单的方法叫做肽指纹谱测定(PMF)[9~13]。这个方法是用特殊的酶对2-DE分离后的蛋白质点进行降解,产生的肽从凝胶中分离出来后再用质谱分析和检测肽的分子量。数据库检测能够产生所有蛋白质理论上的PMF,把它们和质谱分析所获得的肽片进行比较就可以得出结果。第二种方法是在质谱仪中把凝胶分离后的肽分解成片段,产生局部的氨基酸序列(序列标签)。那么就可以用分子量或者序列信息进行数据库检索[14,15]。PMF一般用MALDI-TOF来实现,序列标签可以用串联质谱技术MS-MS进行检测[16~18]。用质谱检测蛋白的灵敏性可以精确到飞摩的水平,甚至已有报道其精确度可以达到阿摩水平[19]。3 用MALDI-TOF进行肽指纹谱分析凝胶分离后的蛋白质鉴定以及肽指纹谱分析是目前质谱实验室常规使用的方法。在进行MALDI分析之前需要最优化的凝胶上消化[20~22]和脱盐过程。胰岛素是最常用的凝胶消化酶,因为它在赖氨酸和精氨酸位点能够特异性切割蛋白质,产生分子量在600~2500Da之间的小分子肽,并能够从凝胶上有效的分离出来。MALDI-TOF是质谱技术中相对简单并很容易使用的一种方式,对样本中的盐和其他污染物有很高的耐受性,这点优于ESI。提取的肽片混合物中较小的片段可以直接沉积在靶板上做PMF分析,剩下的样本可以储存起来作为以后的ESI-MS分析。如果经凝胶分离后所有的肽全用来做MALDI分析,那么样本经脱盐以后将会取得更好的结果。脱盐的过程可以用商品化的试剂盒ZipTip(Millipore),或者在凝胶电泳仪的末端放置一个小的逆向塑料柱来实现[23,24]。PMF中非常重要的一个因素就是质谱测量分子量的准确度[25]。现代的MALDI-TOF仪都具有延迟取样反射器装置,用它们检测的肽段分子量可以精确到10~30ppm。这样的精确度使得4~5个肽段足以清楚的鉴定蛋白质的分子量。MALDI产生的大多数都是单电荷离子,所以它的图谱很容易解释。4 肽质指纹谱分析的自动化为了实现高通量蛋白检测,样本准备,质谱检测,资料分析和数据检索必须实现自动化操作。目前有些实验室用机器人对蛋白质进行自动化取点,凝胶上消化,样本脱盐以及对MALDI平板进行定点测定等。而且,现代化的MALDI-TOF仪完全能够让MALDI检测自动化。资料分析和数据检索过程同样可以自动化。在进行蛋白组分析时,如果在凝胶上不用单个分离就可以对所有蛋白质进行鉴定将是非常具有诱惑力的。为此,研究人员做了很多努力,由Hochestrasser及其同事建立了一套称作分子探测术的方法(molecular scanner)[26,27]。它是将整个2-DE胶上蛋白质样本同时酶解,并将酶解片段一起原位转移至另一张膜上,再直接用MALDI- TOF 质谱对膜上样本进行整体扫描,为实现蛋白质组学研究的自动化、规模化以及临床应用提供了一个崭新的思路和方法。这种思路的优点是显而易见的,不过按照目前的方法,要普及此种技术主要的难点是,质谱扫描一张图谱所需的时间过长,如用0.4mm的精度扫描一张4cm×4cm的膜需55个小时,这就意味着一张16cm×16cm的常用膜的扫描,至少需要36天不间断的工作和大约40G的磁盘空间存储如此庞大的原始数据。5 用ESI-MS/MS的方法创建序列标签只有当被消化的蛋白存在于已有的蛋白或基因组数据库中,并且能从MALDI分析器中得到四五个肽片的数据才能成功进行。如果在EST数据库中只有目的蛋白的部分序列,那么就需要知道肽片的序列信息。创建序列标签的优势在于,用肽的分子量和序列作为数据库检索对象比只用分子量作为检索对象具有更高的特异性。有了序列标签信息后,也可以用EST数据库进行检索[28]。通常来自于一个肽上的序列标签就足以鉴定蛋白质。同MALDI相比,纳升-ESI-MS/MS费时费力,而且在做ESI分析之前必须对样本进行脱盐处理。这些问题可以通过ESI-MS与HPLC联用得到解决。做ESI分析对样本浓度很敏感,用现代化的纳升-HPLC方法可以提供很高的局部肽片凝聚物,分离后的样本可以直接用于质谱分析,而不需要再分解。当分析混合物的时候,在质谱之前做LC分离尤其重要,因为它使得数据依赖的实验(DDE)成为可能,在DDE中,所有的离子都进入检测器进行测量,如果从HPLC到质谱之间出现一个峰的话,软件将自动从质谱转换到MS/MS模式并对产生的离子进行扫描。跟纳升-ESI相比,用这个方法可以从一份标本产生更多的MS/MS数据。Yates等已经详细叙述了如何进行[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]/MS全自动蛋白检测的策略[29]。
[font=宋体]在分子生物学和生物工程领域,蛋白表达载体构建是至关重要的技术之一,它涉及到将目的基因插入到适当的表达载体中,以便在宿主细胞内进行蛋白表达。这一过程不仅有助于研究基因的功能和蛋白质的结构,还为药物开发、基因治疗等领域提供了有力支持。本文将详细介绍蛋白表达载体构建的整个流程,包括目的基因的选择、载体的选择与改造、基因插入、转化、筛选与鉴定等关键步骤。[/font][font=宋体] [/font][b][font=宋体]蛋白表达载体构建流程:[/font][/b][font=宋体] [/font][font=宋体][font=宋体]①[/font][font=Calibri]NCBI[/font][font=宋体]查找目的基因的[/font][font=Calibri]CDS[/font][font=宋体]序列[/font][/font][font=宋体][font=宋体]进入[/font][font=Calibri]NCBI[/font][font=宋体](美国国家生物技术信息中心)的网站。[/font][/font][font=宋体]在搜索框中输入目的基因的名称或关键词。[/font][font=宋体][font=宋体]在搜索结果中找到[/font][font=Calibri]CDS[/font][font=宋体]([/font][font=Calibri]Coding Sequence[/font][font=宋体])序列,通常会显示基因的[/font][font=Calibri]DNA[/font][font=宋体]序列。[/font][/font][font=宋体][font=宋体]记录或复制所需的[/font][font=Calibri]CDS[/font][font=宋体]序列信息。[/font][/font][font=宋体] [/font][font=宋体]②选择合适的表达载体[/font][font=宋体]根据目的基因的性质和所需的表达水平,选择适合的表达载体。[/font][font=宋体]考虑载体的克隆容量、复制子类型、筛选标记等。[/font][font=宋体] [/font][font=宋体]③确定双酶切位点[/font][font=宋体]根据目的基因和载体,选择合适的双酶切位点。[/font][font=宋体][font=宋体]确保酶切位点在[/font][font=Calibri]CDS[/font][font=宋体]序列和载体上都是独特的,避免切割其他部位。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]④[/font][font=Calibri]Primer 5[/font][font=宋体]预测目的序列酶切位点[/font][/font][font=宋体][font=宋体]使用[/font][font=Calibri]Primer 5[/font][font=宋体]或其他相关软件,根据已知的[/font][font=Calibri]CDS[/font][font=宋体]序列设计酶切位点的引物。[/font][/font][font=宋体]通过软件预测引物的特异性,确保它们仅与目的基因结合。[/font][font=宋体] [/font][font=宋体] [/font][font=宋体][font=宋体]⑤双酶切[/font][font=Calibri]buffer[/font][/font][font=宋体][font=宋体]根据选择的酶,准备相应的酶切[/font][font=Calibri]buffer[/font][font=宋体]。[/font][/font][font=宋体][font=宋体]注意[/font][font=Calibri]buffer[/font][font=宋体]的[/font][font=Calibri]pH[/font][font=宋体]值和离子浓度,确保酶的活性。[/font][/font][font=宋体] [/font][font=宋体] [/font][font=宋体][font=宋体]⑥目的基因[/font][font=Calibri]CDS[/font][font=宋体]序列引物设计[/font][/font][font=宋体][font=宋体]根据[/font][font=Calibri]CDS[/font][font=宋体]序列,设计一对引物用于[/font][font=Calibri][url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url][/font][font=宋体]扩增目的基因。[/font][/font][font=宋体]确保引物的特异性,避免与其他基因序列发生非特异性结合。[/font][font=宋体] [/font][font=宋体] [/font][font=宋体]⑦转化[/font][font=宋体][font=宋体]制备感受态细胞,使其处于易于接受外源[/font][font=Calibri]DNA[/font][font=宋体]的状态。[/font][/font][font=宋体][font=宋体]将[/font][font=Calibri][url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url][/font][font=宋体]扩增得到的目的基因与载体混合,进行转化反应。[/font][/font][font=宋体]将转化后的细菌在选择培养基上培养,筛选含有重组载体的阳性克隆。[/font][font=宋体] [/font][font=宋体] [/font][font=宋体]⑧双酶切及鉴定[/font][font=宋体][font=宋体]提取阳性克隆的质粒[/font][font=Calibri]DNA[/font][font=宋体]。[/font][/font][font=宋体]使用设计的引物进行双酶切反应。[/font][font=宋体]对酶切产物进行电泳分析,观察是否获得预期的片段,并进行凝胶回收。[/font][font=宋体] [/font][font=宋体] [/font][font=宋体]⑨测序[/font][font=宋体]将回收的酶切产物进行序列测定。[/font][font=宋体][font=宋体]对比测序结果与目的基因的[/font][font=Calibri]CDS[/font][font=宋体]序列,确保没有突变或错误。[/font][/font][font=宋体] [/font][font=宋体] [/font][font=宋体]⑩阳性菌落扩大培养,用于后期蛋白纯化研究[/font][font=宋体] [/font][font=宋体]选择测序正确的阳性菌落进行扩大培养。[/font][font=宋体]在摇瓶或发酵罐中进行高密度培养,为后续的蛋白表达提供充足的原料。[/font][font=宋体]在确定目的基因在载体上正确表达后,可以进行蛋白纯化研究。[/font][font=宋体]通过适当的纯化技术,如亲和层析、离子交换等,分离和纯化目的蛋白。[/font][font=宋体]对纯化的蛋白进行质量分析和功能研究。[/font][font=宋体] [/font][font=宋体][font=宋体]综上所述,蛋白表达载体构建是一个系统性的过程,涉及到多个关键步骤和复杂的技术操作。通过遵循这一流程,研究人员能够成功地在宿主细胞内表达所需蛋白,并进一步对其进行分析和功能研究。随着生物技术的不断发展,蛋白表达载体构建的应用领域将越来越广泛,为人类对生命现象的深入理解和疾病的防治提供更多可能性。因此,不断优化和完善这一技术对于生命科学领域的发展至关重要。更多[url=https://cn.sinobiological.com/resource/protein-review/protein-production][b]蛋白表达生产[/b][/url]详情可以关注[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/protein-production[/font][/font]
蛋白质组学研究的一般工具与方法随着人类基因组计划取得巨大的成功和许多物种基因组测序的完成,仅仅靠基因组的序列来试图阐明生命现象是远远不够的,因此,研究重心已经开始从揭示生命的所有遗传信息转移到在分子整体水平对功能的研究上,生命科学已实质性地跨入了后基因组时代。 尽管现在已经有多个物种的基因组被测序,但这些基因组中通常有一半以上基因的功能是未知的。目前功能基因组研究中所采用的策略,如微阵列法(microarray)(Wodicka et al., 1997)、基因芯片(gene chips)(Ramsay et al., 1998)、基因表达序列分析(SAGE)(Velculescu et al., 1995)等,都是从细胞中mRNA的角度来考虑的。但事实上,从DNA、mRNA到蛋白质存在三个层次的调控,mRNA自身也存在着贮存、转运和降解等问题,从mRNA角度考虑,实际上仅包括了转录水平调控,并不能全面代表蛋白质表达水平。实验也证明,组织中mRNA丰度与蛋白质丰度的相关性并不好,尤其对于低丰度蛋白质来说,相关性更差。蛋白质复杂的翻译后修饰,蛋白质的亚细胞定位或迁移,蛋白质-蛋白质相互作用则几乎无法从mRNA水平来判断(曾嵘,夏其昌,2002)。新生肽链合成后存在多种加工、修饰过程,蛋白质间也存在类似于mRNA分子内的剪切、拼接,研究证明基本元件“intein”广泛存在于蛋白质中(Perler et al., 1997)。基因与其编码产物蛋白的线性对应关系只存在于新生肽链而不是最终的功能蛋白质中。 蛋白质是生理功能的执行者和生命现象的直接体现者,对蛋白质结构和功能的研究将直接阐明生命在生理或病理条件下的变化机制;蛋白质本身的存在形式和活动规律,如翻译后修饰、蛋白质间相互作用及蛋白质构象等问题,仍依赖于直接对蛋白质的研究来解决。因此要对生命的复杂活动有全面和深入的认识,必然要在整体、动态、网络的水平上对蛋白质进行研究(钱小红,贺福初,2003)。 蛋白质组学研究中常用的技术体系 方法学上,二维凝胶电泳-质谱仍然是目前最流行和可靠的技术平台(Rabilloud et al., 2000)。其一般过程是:细胞或组织样品——样品制备——二维凝胶电泳(2D-PAGE)分离蛋白质——计算机辅助分析2D-PAGE图象——对感兴趣的蛋白质进行酶解——质谱分析——数据库检索——蛋白质鉴定——分析蛋白质在细胞与组织中的表达情况。 2-D PAGE 样品制备 2D-PAGE 的操作流程基本上实现了程序化。但是,样品制备是一个非常关键与复杂的过程。成功的2D-PAGE取决于对样品中蛋白质有效的抽提和它的溶解性。与核酸不同,目前没有一种通用的方法适用于所有的蛋白质,来源不同的蛋白质都受到自身蛋白质制备方法的挑战。 正确的样品制备方法从收集样品开始时就要防止样品的裂解和被蛋白水解酶降解(Rabilloud et al., 2000)。要尽可能溶解更多的蛋白,并且在2D-PAGE过程中保持它的溶解性,阻止蛋白质的人为修饰。在样品制备过程中,各个实验室也通过实验建立了更为可行的方法。目前通过建立分步提取方法可以有效地提取出更多的蛋白质(兰彦等,2001)。另一种对蛋白质采用预分离的方法称为“多间隔电解法(multi-compartment-electrolyser)”,采用这种方法后,分辨率和胶的质量均明显改善(Herbert et al., 2000)。 但是,由于生物样品的多样性和复杂性,目前所采用的样品制备方法具有局限性。其它物质对蛋白质样品制备存在干扰。核酸通过与蛋白质结合,增加样品黏度而干扰等点聚焦(IEF)分离的效果。当然,通过实验探索,采取一些措施可以减轻它的干扰。例如,在样品制备过程中加入非特异性的核酸酶或RNase与DNase的混合物,在等电聚焦时将每个胶条的电流限制在50mA以内通常可以消除其影响。脂类物质的影响可以通过利用有机溶剂的方法将其去除,但是这常常会导致蛋白质的不可逆沉淀。除了蛋白质的降解之外,糖基化是蛋白质的最重要的人工修饰,样品中的尿素在这一过程中起着非常重要的作用。样品中的尿素在降解的过程中会形成能够与蛋白质的氨基反应的氰酸盐,这种结果会导致蛋白质带有更多的正电荷。所以,在2D-PAGE中要用新鲜的尿素溶液,在等电聚焦过程中要控制温度不能太高(Beranova-Giorgianni, 2003)。但是,目前还没有一种简单有效的方法来去除样品中的多糖。 样品分离和分析 样品制备完成后运用IEF和SDS-PAGE电泳对它进行分离,常采用银染和考马斯亮兰染色即可观察到具有许多蛋白质斑点的凝胶图像。等电聚焦电泳与SDS-PAGE的具体操作步骤已经实现了程序化,均有详细操作流程参考,但是由于样品的不同,不同样品的具体条件还需要试验探索。第二相SDS-PAGE运行结束,染色完毕后,利用计算机软件对凝胶图像进行分析,如PD-QUEST软件,LIPS,HERMES,GEMINI等,对凝胶图像上的蛋白质斑点进行匹配,对图像进行数字化处理等分析(贾宇峰等,2001),对感兴趣的蛋白质采用质谱分析。 低丰度蛋白质的检测 低丰度蛋白在蛋白质组学研究中常常是人们非常感兴趣的,因为细胞或组织中的一些生物活性物质,如细胞分泌的一些活性物质,受体等表达量都非常低。按照一般电泳的上样量,这些小分子是根本看不到的,但如果单纯地增加上样量,细胞或组织中的大量表达的蛋白就会将其覆盖,而且上样量过大也会影响电泳结果。所以对这些低丰度的样品可以进行富集,富集的方法可以通过层析,如亲和层析,离子交换层析等方法,还可以通过利用样品等电点性质等方法将pH范围相近的蛋白质富集(Santoni et al., 2000; Beranova-Giorgianni, 2003)。
目前,违法添加非食用物质和滥用食品添加剂事件常有发生,为保证牛奶的食用安全,采用离子色谱仪检测牛奶中硫氰酸根。 1、检测原理:液态奶样品沉淀蛋白、去除脂肪后,用离子色谱分析,电导检测器检测,外标法定量。 2、实验部分 2.1试剂与材料 2.1.1试验用水均为超纯水 2.1.2乙腈(色谱纯) 2.1.3固相萃取小柱:OnGuardRP柱(2.5cc),或相当者(如C18),使用前依次用5ml甲醇和10ml水活化。 2.1.4硫氰酸标准品:北京化工厂 2.1.5硫氰酸标准储备液将硫氰酸标准品于80度烘箱内烘干2小时。准确称取干燥后的硫氰化钾1.6732g于1000ml容量瓶中,定容,混匀。即得1000ppm硫氰根标准储备液。 2.1.6硫氰酸标准中间液取硫氰酸标准储备溶液1mL,置于100mL容量瓶中,加水至刻度。此溶液含硫氰酸10mg/L。 2.1.7硫氰酸标准使用液移取0.1、0.2、0.5、1.0、2.0mL硫氰酸标准中间液,用水定容于10mL容量瓶中,浓度分别为0.1、0.2、0.5、1.0、2.0mg/L。 2.2仪器 2.2.1离子色谱仪:配备淋洗液发生器和电导检测器; 2.2.2离心机:冷冻离心机。 2.3样品处理取4mL液体奶样品,加入5mL乙腈沉淀蛋白,取上清液稀释10倍,过RP柱(或经冷冻离心机)去除脂肪后上机。 2.4离子色谱参考条件色谱柱:强亲水性阴离子交换柱。IonPacAS16,4.0×250mm分析柱;IonPacAG16,4.0×50mm保护柱;或其他相当者。 采用离子色谱仪检测,操作简便,检测结果准确。
[b][font=宋体]整合蛋白和跨膜蛋白定义:[/font][/b][font=宋体] [/font][font=宋体]整合蛋白和跨膜蛋白是两类重要的蛋白质,它们在细胞分子水平上起着重要的作用。[/font][font=宋体] [/font][font=宋体]整合蛋白,也称为内在蛋白或跨膜蛋白,部分或全部镶嵌在细胞膜中或内外两侧,以非极性氨基酸与脂双分子层的非极性疏水区相互作用而结合在质膜上。它们是生物膜的基本结构成分,许多具重要生理功能的膜蛋白均属整合蛋白,如膜结合的酶类、载体蛋白、通道蛋白、膜受体等。[/font][font=宋体] [/font][font=宋体]跨膜蛋白,是可以跨越细胞膜的蛋白,它在细胞的信号传递系统中担当着重要的角色。跨膜蛋白在结构上可以分为单次跨膜、多次跨膜、多亚基跨膜等,它们具有能够跨越细胞膜的能力。[/font][font=宋体] [/font][b][font=宋体]整合蛋白和跨膜蛋白在位置、结构和功能上存在显著的差异[/font][/b][font=宋体] [/font][font=宋体]①位置:整合蛋白主要存在于细胞质内,细胞核或其他非细胞膜结构中,它们容易在细胞中自由移动。而跨膜蛋白则嵌入细胞膜中,一部分位于细胞膜的胞外侧,另一部分位于细胞膜的胞内侧,形成了一个穿过细胞膜的通道。[/font][font=宋体][font=宋体]②结构:整合蛋白的结构通常由两个独立的部分组成,一个是靠近细胞膜的膜结合区域([/font][font=Calibri]TM[/font][font=宋体]),另一个是靠近细胞骨架的非膜结合区域([/font][font=Calibri]N-TM[/font][font=宋体])。当接受到外界的信号时,整合蛋白的[/font][font=Calibri]TM[/font][font=宋体]区域会被激活,把来自外界的信号转化为细胞内可以识别的信号,直接参与细胞信号传导系统中。[/font][/font][font=宋体]③功能:整合蛋白主要是用来从外界传达信号到细胞内,充当细胞与外界信号的桥梁。而跨膜蛋白则在细胞的信号传递系统中担当着重要的角色。[/font][font=宋体]总的来说,整合蛋白和跨膜蛋白在位置、结构和功能上存在显著的差异,这些差异使得它们在生物体中扮演着不同的角色。[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/resource/protein-review/transmembrane-proteins][b]跨膜蛋白表达与制备服务[/b][/url],制备流程图:基因合成[/font][font=宋体]→载体构建→细胞转化[/font][font=Calibri]/[/font][font=宋体]转染→蛋白表达→细胞收集→细胞破碎→膜脂提取→膜脂增溶→蛋白纯化→质量检测,同时义翘拥有[/font][/font][b][font=宋体]三大跨膜蛋白制备平台[/font][/b][font=宋体],可以为客户提供全面的多次跨膜蛋白产品和服务。同时,为基础研究和药物研发提供更加优质的原材料。[/font][font=宋体] [/font][b][font=宋体][font=Calibri]VLP[/font][font=宋体]技术平台[/font][/font][/b][font=宋体][font=宋体]正确折叠的膜蛋白在细胞膜上表达,类病毒颗粒[/font][font=Calibri]VLP[/font][font=宋体]通过出芽的方式包裹上携带有靶标蛋白的细胞膜,形成包膜的[/font][font=Calibri]VLP[/font][font=宋体]。它是由病毒的衣壳蛋白通过自组装而形成的纳米级颗粒(直径约[/font][font=Calibri]100[/font][font=宋体]~[/font][font=Calibri]300[/font][font=宋体]纳米),不含病毒核酸,不能进行自主复制,生产操作过程中较为安全。产生的[/font][font=Calibri]VLP[/font][font=宋体]蛋白可直接像可溶蛋白一样进行包被进行[/font][font=Calibri]ELISA[/font][font=宋体]检测。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州已成功开发[/font][font=Calibri]VLP[/font][font=宋体]技术平台,它可以将完整天然构象的膜蛋白展示在类病毒颗粒表面,这种方法不仅可以保留膜蛋白的完整结构,同时也能够真实地模拟其在细胞膜上的位置和构象。[/font][/font][font=宋体] [/font][b][font=宋体]去垢剂技术平台[/font][/b][font=宋体][font=宋体]由于存在疏水结构域,跨膜蛋白与膜的结合非常紧密,需要用去垢剂([/font][font=Calibri]detergent[/font][font=宋体])才能从膜上洗涤下来,[/font][font=Calibri]Detergent[/font][font=宋体]作为一种两亲性分子,疏水尾部包裹目的蛋白的疏水区域,亲水头部位于与溶液接触的界面。微团的形成是膜蛋白增溶的基础,当去垢剂浓度高于[/font][font=Calibri]CMC[/font][font=宋体]([/font][font=Calibri]Critical micelle concentration[/font][font=宋体],临界胶束浓度)时会形成微团,增溶后,去垢剂将蛋白周围的磷脂置换,从而实现收集目标膜蛋白的目的,后续再进行蛋白纯化,最终蛋白呈现在含有[/font][font=Calibri]Detergent[/font][font=宋体]的溶液中。义翘神州成功搭建了去垢剂技术平台,利用该平台可有效提高跨膜蛋白的产量和纯度。[/font][/font][font=宋体] [/font][b][font=宋体][font=Calibri]Nanodisc[/font][font=宋体]技术平台[/font][/font][/b][font=宋体][font=Calibri]Nanodisc[/font][font=宋体]结构稳定,与天然的生物膜非常相似,使得[/font][font=Calibri]Nanodisc[/font][font=宋体]能够很好地应用于膜蛋白的研究。目前[/font][font=Calibri]Nanodisc[/font][font=宋体]平台有[/font][font=Calibri]2[/font][font=宋体]种方式,一种是基于苯乙烯马来酸酐共聚物([/font][font=Calibri]SMA[/font][font=宋体])组装的[/font][font=Calibri]SMA-Nanodisc[/font][font=宋体]平台,如下图(左)所示,它可以直接从细胞膜上提取膜蛋白,使其变为可溶性蛋白,组装完成的蛋白样品很稳定,更能维持蛋白的天然构象。另一种是基于膜骨架蛋白([/font][font=Calibri]MSP[/font][font=宋体])的[/font][font=Calibri]MSP-Nanodisc[/font][font=宋体]平台(下图右),它需要先将膜蛋白利用去垢剂制备出来,然后再加入磷脂分子和[/font][font=Calibri]MSP[/font][font=宋体]进行组装。通过调整磷脂、[/font][font=Calibri]MSP[/font][font=宋体]和待组装膜蛋白三者的比例,可以使得待组装膜蛋白在[/font][font=Calibri]Nanodisc[/font][font=宋体]中呈不同聚集状态。义翘神州已成功搭建了[/font][font=Calibri]Nanodisc[/font][font=宋体]技术平台,利用跨膜蛋白与磷脂结合能够维持其良好活性的特性,制备出稳定的产品,满足动物免疫、抗体筛选、[/font][font=Calibri]cell-based assays[/font][font=宋体]等场景。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/transmembrane-proteins[/font][/font]







 我要推广仪器
我要推广仪器
 下载APP
下载APP