[font=宋体][font=宋体]无细胞蛋白表达是一种体外重组蛋白质表达技术也称为无细胞蛋白质合成技术([/font][font=Calibri]CFPS[/font][font=宋体]:[/font][font=Calibri]Cell-free protein synthesis[/font][font=宋体]),是指用含有蛋白合成必需的组分(核糖体,转运[/font][font=Calibri]RNA[/font][font=宋体],氨酰合成酶,启动[/font][font=Calibri]/[/font][font=宋体]延伸[/font][font=Calibri]/[/font][font=宋体]终止因子,三磷酸鸟苷,[/font][font=Calibri]ATP[/font][font=宋体],[/font][font=Calibri]Mg2+[/font][font=宋体]和[/font][font=Calibri]K+[/font][font=宋体])的细胞裂解物在体外进行蛋白合成。无细胞蛋白表达技术适用于制备各种类型的蛋白质,包括难表达蛋白质、毒性蛋白质、复杂蛋白质等。在药物研究、生物制造和生命科学等领域中得到广泛关注和应用,无论是研究、开发还是商业化应用过程。目前无细胞蛋白表达主要应用于药物研发领域,例如抗体制备和生物药物生产等。随着人工智能技术的不断发展,无细胞蛋白表达技术可以与人工智能算法结合,构建计算机辅助的高通量生产系统,实现个性化、精准的生物医学治疗。除此之外,还能够应用于其他领域,例如基因工程、环境保护和农业生产等。随着无细胞蛋白表达技术的不断发展和人工智能技术的不断进步,我们可以看到更多的新领域和新应用出现,给生物科技行业带来更多的机遇和挑战。[/font][/font][font=宋体] [/font][font=宋体] [/font][b][font=宋体]相较于传统的活细胞蛋白表达技术,无细胞蛋白表达技术具有以下几个显著的优势:[/font][/b][font=宋体] [/font][font=宋体][font=Calibri]1. [/font][font=宋体]更高的蛋白质表达量:传统的活细胞蛋白表达技术受限于细胞本身的多方面因素,其表达的蛋白质数量往往受到限制。而无细胞蛋白表达技术通过在体外底物浓度高的环境中进行合成反应,不但避免了传统活细胞表达所面临的方方面面的限制,还能够很好地控制反应体系,从而获得表达量更高的蛋白质。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2. [/font][font=宋体]更快的表达速度:传统活细胞蛋白表达需要细胞生长并达到最佳密度才能进行蛋白质表达,这个过程往往需要数天时间。而无细胞蛋白表达技术通常只需要数小时就能够完成蛋白质的表达,这个速度明显快于传统活细胞表达技术。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3. [/font][font=宋体]更精准的蛋白质合成:无细胞蛋白表达技术在体外进行蛋白质合成,能够精确控制底物浓度、反应温度、反应剂比例等参数,因此可以更加精准地合成定制的蛋白质,这对于研究和应用来讲具有重要意义。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4. [/font][font=宋体]更灵活控制:在无细胞蛋白表达技术中,可以使用分离的组分体系进行蛋白质的合成,可以控制底物和反应剂的比例,也可以在适当的反应条件下进行自定义的修饰,如蛋白质标记、药效分析等。这些优点使得无细胞蛋白表达技术更加灵活、可控,适用于更广泛的应用领域。[/font][/font][font=宋体] [/font][b][font=宋体] [/font][font=宋体]无细胞蛋白表达应用[/font][/b][font=宋体] [/font][font=宋体]无细胞蛋白表达技术是一种飞速发展的新型生物技术,具有广阔的应用前景和潜力。该技术可以快速、高效、经济地合成蛋白质,可广泛应用于医疗、制药、农业、生物材料等多个领域。[/font][font=宋体] [/font][font=宋体][font=Calibri]1. [/font][font=宋体]医疗领域:无细胞蛋白表达技术在医疗领域应用广泛,可以用于生产多种蛋白质药品,如单克隆抗体等。其中,单克隆抗体是一种重要的治疗药物,具有高度特异性和亲和力,可用于肿瘤、心血管疾病、自身免疫性疾病等疾病的治疗。传统单克隆抗体生产方法需要花费大量时间和成本,而无细胞蛋白表达技术则可以在短时间内大规模合成单克隆抗体,从而大大缩短生产周期,并且可以降低成本。此外,无细胞蛋白表达技术也可以用于疫苗研发。比如疟疾疫苗研究开发昂贵又耗时,目前利用[/font][font=Calibri]WGE[/font][font=宋体]系统可加速疫苗研发,并建立高通量疟原虫抗体筛查系统。[/font][font=Calibri]Stark[/font][font=宋体]等利用大肠杆菌的便携式冻干裂解物再水化,[/font][font=Calibri]1h[/font][font=宋体]内合成高致病性病原体土拉弗朗西斯菌亚种的生物偶联疫苗,与工程菌生产的疫苗相比,其可引发更高水平的病原体特异性抗体。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2. [/font][font=宋体]制药领域:是无细胞蛋白表达技术的一个重要应用领域。药物开发的成功率取决于药物分子对目标蛋白的亲和力,而目标蛋白对于专一的细胞表达系统和分类的组织或器官非常敏感。通过无细胞蛋白表达技术,研究人员可以在不依赖于细胞的情况下直接生产大量需要的蛋白质,为药物研发提供了更快更便捷的方法。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3. [/font][font=宋体]基础研究领域:利用无细胞蛋白质合成系统可以直接对表达产物进行核磁共振分析,目前已确定了数千个蛋白质的结构。可以通过合成蛋白质建立蛋白质阵列,解开基因产物的功能;应用核糖体展示和 [/font][font=Calibri]mRNA [/font][font=宋体]展示技术,更有利于实现高通量筛选,全面深入研究基因特征和功能。通过无细胞蛋白表达技术可以实现对大型蛋白质的生产和分析,同时也为基础研究打开了新的研究领域。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]目前义翘神州无细胞合成服务正在活动中,活动时间[/font][font=Calibri]2023[/font][font=宋体]年[/font][font=Calibri]10[/font][font=宋体]月[/font][font=Calibri]23[/font][font=宋体]日[/font][font=Calibri]-12[/font][font=宋体]月[/font][font=Calibri]31[/font][font=宋体]日。有需求的可以咨询或者进入义翘神州网进行查看。更多详情可以关注[/font][font=Calibri]https://cn.sinobiological.com/services/cell-free-protein-synthesis-service[/font][/font]
细胞的癌变是细胞在信号通路调节失控情况下的无限制增生,而RAF-MEK-ERK信号通路的持续性激活则是诱导细胞癌变的重要原因。因此,对RAF-MEK-ERK信号通路的研究一直是分子生物学研究的热点。英国科学家在最近一期《分子与细胞生物学》杂志上发表论文称,真核翻译起始因子3a(EIF3a)可以通过和RAF激酶结合,抑制RAF-MEK-ERK信号通路,是这一信号通路的重要“刹车”蛋白。这一发现意味着EIF3a可能成为下一代抗癌药物全新的靶标蛋白,为抗癌药物的研发提供新思路。 EIF3a是细胞蛋白质翻译起始复合物的重要构件。由英国格拉斯哥大学和爱尔兰都柏林大学研究人员组成的研究小组研究发现,EIF3a能够与细胞外信号调节激酶通路的两个组成部分SHC蛋白和Raf1蛋白绑定,不仅可以调节蛋白质翻译,影响细胞的生长和分化,还通过和RAF激酶结合,抑制RAF-MEK-ERK信号通路,成为RAF-MEK-ERK信号通路的重要“刹车”蛋白。同时,研究人员还发现,EIF3a和另一个“刹车”蛋白——β抑制蛋白(β-arrestin2)结合,能够调节细胞的另一条最主要信号通路:G蛋白偶联受体(GPCR)信号通路。 该论文首席作者,格拉斯哥大学生物医学与生命科学学院的徐天瑞博士指出, RAF-MEK-ERK信号通路是细胞外信号传递入细胞内的主干通路,绝大多数细胞外信号都可以通过RAF-MEK-ERK通路影响细胞行为,诸如细胞增值、细胞分化、细胞凋亡。新发现表明,EIF3a可以抑制RAF-MEK-ERK信号通路,从而抑制癌症的产生;其通过与β抑制蛋白结合影响其功能,可治疗由G蛋白偶联受体信号通路失控而导致的癌症;同时,EIF3a可以抑制RAF激酶活性,诱导细胞凋亡,从而杀灭癌细胞;而对EIF3a本身蛋白质翻译的调节,也可以作为治疗癌症的重要手段。 徐天瑞博士说:“信号抑制蛋白EIF3A的发现意义重大,《科学—信号传导》杂志最近将其列为2011年度细胞生物学领域八项重大进展之一。我们的新研究证明,EIF3a不仅是蛋白质翻译起始因子,也是RAF-MEK-ERK信号通路和G蛋白偶联受体信号通路交汇点上的重要调控蛋白。通过对EIF3a的调控,有可能四管齐下地杀灭癌细胞,从而使EIF3a可能成为下一代抗癌药物全新的靶标蛋白,为抗癌药物的研发提供新思路。”(记者 刘海英)
[b][font=宋体]前言[/font][/b][font=宋体]在蛋白质研究领域,稳定细胞系的应用已成为生产高质量结构生物学蛋白质的关键手段。随着技术的不断进步,稳定细胞系的生成与筛选方法得到了显著改进,从而推动了蛋白质生产的高效化与精准化。[/font][font=Calibri] [/font][b][font=宋体]细胞系的建立和应用[/font][font=宋体][font=Calibri]HEK293[/font][font=宋体]和[/font][font=Calibri]CHO[/font][font=宋体]细胞系[/font][/font][/b][font=宋体]因其稳定的蛋白表达和适当的翻译后修饰而被广泛用于结构生物学研究。这些细胞系能有效地生产具有复杂糖基化模式的蛋白质,这对于确保蛋白质的功能和稳定性至关重要。糖基化缺陷细胞系通过特定的基因改造,能够分泌脱糖基化糖蛋白,为蛋白质生产提供了更加纯净的原料。[/font][font=Calibri] [/font][b][font=宋体]稳定细胞系的生成[/font][/b][font=宋体][font=宋体]传统的稳定细胞系生成技术如瞬时转染,虽然方法简便,但存在整合频率低、转基因沉默等问题。为了克服这些困难,研究者们开发出了一系列新技术,如细胞分选技术、位点特异性重组(如[/font][font=Calibri]FLP/FRT[/font][font=宋体]系统)、转座子系统(如[/font][font=Calibri]piggyBac[/font][font=宋体])、慢病毒系统以及噬菌体整合酶等,提高了稳定细胞系的生成效率和稳定性。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]序列特异性基因组工程也为稳定细胞系的生成提供了新的思路。通过敲除或修饰特定的基因,研究者们能够实现对细胞功能的精准调控,从而优化蛋白质生产的效率和纯度。例如,一种同时缺乏[/font][font=Calibri]GnTI[/font][font=宋体]和谷氨酰胺合成酶([/font][font=Calibri]GS[/font][font=宋体])活性的[/font][font=Calibri]CHO[/font][font=宋体]细胞系被成功开发出来,为高效筛选具有[/font][font=Calibri]GS[/font][font=宋体]标记的稳定细胞系提供了有力工具。[/font][/font][font=Calibri] [/font][b][font=宋体]稳定细胞系与瞬时转染的比较[/font][/b][font=宋体]稳定细胞系相较于瞬时转染具有多个优点,包括能够进行大规模生产和保持高水平的蛋白表达稳定性。尽管瞬时转染在某些情况下能快速产生大量蛋白,但其表达水平和重复性通常不如稳定细胞系。[/font][font=Calibri] [/font][b][font=宋体]展望[/font][/b][font=宋体]近年来,利用稳定细胞系高效生产结构生物学蛋白质已成为研究的热点和趋势。通过引入新技术、优化筛选方法和改进整合系统,不仅能够提高蛋白质生产的效率和纯度,还能够为结构生物学研究提供更加精准、可靠的实验工具。随着基因编辑和细胞工程技术的进步,预计在未来,通过精确的基因操作能够更有效地创建和利用稳定细胞系。这些技术的进步将促进结构生物学和药物开发中蛋白质的高效和可持续生产。[/font][font=宋体] [/font][font=宋体]本文由义翘神州进行整理,同时提供[/font][url=https://cn.sinobiological.com/services/stable-cell-line-development-service][u][font=宋体][color=#0000ff]稳定细胞系构建服务[/color][/font][/u][/url][font=宋体],详情可点击了解![/font][font=Calibri] [/font][font=宋体]参考文献:[/font][font=Calibri]Büssow K. Stable mammalian producer cell lines for structural biology. [/font][i][font=Calibri]Curr Opin Struct Biol[/font][/i][font=Calibri]. 2015 32:81-90. doi:10.1016/j.sbi.2015.03.002[/font]
[font=宋体][font=宋体]无细胞蛋白表达系统([/font][font=Calibri]Cell-Free Protein Expression System[/font][font=宋体])是一种基于原核和真核细胞提取物构建的体外蛋白表达系统。它具有许多优点,例如可以在短时间内生产大量的蛋白质,同时避免了细胞内的复杂调控机制和翻译后修饰等繁琐过程。因此,无细胞蛋白表达系统在生物制药、生物材料、生物燃料等领域具有广泛的应用前景。本文将详细介绍无细胞蛋白表达系统的优缺点。[/font][/font][font=宋体][b]一、无细胞蛋白表达系统的优点[/b][/font][font=宋体] [/font][font=宋体][font=Calibri]1.[/font][font=宋体]高效性:无细胞蛋白表达系统具有高表达效率的优点,这是由于体外体系中不存在靶蛋白累积所需的细胞分裂和细胞复杂代谢反应。此外,由于无细胞蛋白表达系统不受到细胞毒性和免疫反应的限制,可以实现大规模的蛋白质表达。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2.[/font][font=宋体]灵活性:无细胞蛋白表达系统可以使用一系列不同的原核和真核细胞提取物作为反应体系,例如[/font][font=Calibri]E.coli[/font][font=宋体]、小麦胚芽和人类细胞等。这意味着可以根据不同的实验目的和需求进行合理的选择,以适应多样化的研究需要。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3.[/font][font=宋体]易操作性:无细胞蛋白表达系统非常容易操作。与传统的细胞表达系统相比,无细胞蛋白表达系统不需要细胞培养、生长和繁殖。此外,无细胞蛋白表达系统可以快速进行,通常只需要数小时至几天即可完成目标蛋白的表达。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4.[/font][font=宋体]简单纯化:由于无细胞蛋白表达系统可以避免有机溶剂和离子交换剂等复杂的步骤,从而使目标蛋白的纯化工作更加简便和迅速。例如,可以使用亲和柱、凝胶过滤和电泳分析等方法来快速分离和纯化蛋白质。[/font][/font][font=宋体] [/font][font=宋体][b]二、无细胞蛋白表达系统的缺点[/b][/font][font=宋体] [/font][font=宋体][font=Calibri]1.[/font][font=宋体]成本较高:尽管无细胞蛋白表达系统可以大规模进行蛋白质表达,但是所需的原核和真核细胞提取物通常需要较高的成本。此外,涉及到的一些试剂和设备也比较昂贵,使得无细胞蛋白表达系统在应用过程中存在一定的经济压力。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2.[/font][font=宋体]表达限制:由于无细胞蛋白表达系统缺乏复杂的代谢反应和细胞分化机制,因此它不适用于某些特定类型的蛋白。例如,它无法表达复杂的膜蛋白和困难的药物蛋白等。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3.[/font][font=宋体]不稳定性:无细胞蛋白表达系统通常具有一定的稳定性问题。由于缺乏细胞膜的保护,无细胞蛋白表达体系会更容易受到外部条件的影响,如温度、[/font][font=Calibri]pH[/font][font=宋体]、离子浓度等,从而导致蛋白质的不稳定性、聚集和降解等现象。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4.[/font][font=宋体]不适合复杂蛋白结构:无细胞蛋白表达系统对于复杂蛋白结构的模拟效果不佳。例如,膜蛋白、多肽和糖蛋白等复杂蛋白质可能会被无细胞蛋白表达系统无法很好地复制,从而限制了其应用范围。[/font][/font][font=宋体] [/font][font=宋体]无细胞蛋白表达系统具有高效、灵活、易操作、简单纯化等优点,但同时也存在着成本较高、表达限制、不稳定性和不适合复杂蛋白结构等缺点。在实际应用中,需要根据具体的研究目的和需求进行选择,并结合其他技术手段来弥补其不足之处。[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/services/cell-free-protein-synthesis-service][b]无细胞蛋白表达服务[/b][/url],服务优势:[/font][font=宋体]①快速、高效 ②高成功率 ③一致性 ④高难度抗体表达[/font][/font][font=宋体] [/font][font=宋体][font=宋体]详情可以咨询,具体[/font][font=Calibri]https://cn.sinobiological.com/services/cell-free-protein-synthesis-service[/font][/font]
我们知道,PeproTech的所有细胞因子和蛋白均为冻干粉,这使得运输非常便捷,只要常温即可。而且,细胞因子和蛋白冻干粉非常稳定,在-20℃或-80℃条件下可保存数年。冻干粉在使用前需进行溶解,然后以液体形式加到培养体系或注射入动物体内。溶解步骤非常关键,因溶解不好会导致细胞因子或蛋白的失活,这也是很多用户在实际使用中经常遇到的问题。 那么,应该如何进行正确的溶解呢? 下面我们以Recombinant Human IL-4 (重组人IL-4,产品编号:200-04)的说明书为例,对细胞因子或蛋白的溶解方法进行详细的阐述。 拿到重组人IL-4的说明书后,您会发现有一段关于Reconstitution(重悬)的叙述,这段内容含有溶解相关的所有信息。 1. Centrifuge the vial prior to opening.第1步:开盖前离心试剂管 PeproTech的细胞因子或蛋白冻干粉装盛在塑料管中,为无菌包装。冻干粉在运输过程中可能会因颠簸而漂散并粘贴于管壁或管盖上,所以在打开塑料瓶盖前,需将冻干粉通过离心收集到管底,以便用很小体积的液体即可将冻干粉完全溶解。 有很多用户会问一个问题,即应该用多少转速、多长时间离心试剂管,才能达到良好的收集效果?答:有些小型高速离心机(多为进口品牌)的面板上有一个Spin键,按了此键后,离心机会自动快速上升到其最大速度(10000rpm或12000rpm),上升到最高点后速度即刻下降,直至停止旋转,整个过程大约30s。这个Spin键足以很好的将细胞因子或蛋白收集到管底。 但有些实验室没有这样的高速离心机,只有最高转速为4000-4500rpm的离心机。这种情况下,需3000-3500rpm离心5min,也能达到类似的效果。 2. Reconstitute in water to a concentration of 0.1-1.0 mg/ml. Do not vortex.第2步:用无菌水重悬至0.1-1.0 mg/ml,不可振荡。 这个步骤即为溶解步骤,非常重要。 1) 一定要用推荐的溶液重悬(或溶解)冻干粉 用于溶解细胞因子或蛋白的溶液千差万别。此例中的重组人IL-4需用水溶解,而重组人IL-2 (产品编号:200-02)则需用100mM Acetic Acid (醋酸)溶解,重组人TGF-beta1 (产品编号:100-21)需用10mMCitric Acid (柠檬酸),pH3.0溶解,重组人FGF-basic(产品编号:100-18B)需用5mMTris,pH7.6溶解,重组人FGF-10(产品编号:100-26)需用5mMSodium Phosphate(磷酸钠),pH7.4溶解,重组IL-13(产品编号:200-13)需用20mMMHCl溶解。(注:即使同一重组细胞因子或蛋白,不同批次的溶解方法也可能有所不同,因此上面的叙述仅供参考。具体应该如何溶解应以相应批次的官方说明书为准)。后续的文章中将对各种溶解液的配制方法进行详细的阐述,望继续关注。 经常有用户会问,为什么会有这么多种溶解方法?答:我们知道,蛋白的溶解性与很多因素有关,其中比较重要的是pH值和离子强度。PeproTech的细胞因子或重组蛋白在出厂前均经严格测试,说明上所标明的溶解液是能够将该细胞因子或重组蛋白完全溶解的液体。如果您所用的溶解液的pH值和离子强度与说明书中所标明的不符,很多时候会造成细胞因子或重组蛋白不能完全溶解或者根本无法溶解,这样所配得的细胞因子或重组蛋白必然活性不够或丧失。 有不少用户没注意说明书上的描述,而是根据习惯,直接用PBS或培养液(1640或DMEM等)等溶解细胞因子或蛋白的冻干粉。这样做可以吗?答:有时可以,要看具体情况。PeproTech的大多数细胞因子或蛋白冻干粉的溶解液不是PBS,此时千万不能用PBS或培养液直接来溶解,具体原因在上面已经叙述过。而有部分细胞因子,如重组人KGF(产品编号:100-19)和重组人FGF-23(产品编号:100-52)等,说明书上的溶解液即为1x PBS,此时用PBS溶解完全没问题,那么用培养液溶解也是可以的,不过最好还是用先用PBS溶解,然后再用培养液稀释。
中国科技网伦敦9月20日电 英国科学家在最近一期《临床调查杂志》上刊发论文称,他们利用新的检测技术证明,导致亨廷顿病的有害蛋白是逐渐在血液细胞中积累起来的。他们在论文中对这些有害细胞是如何损害人的大脑进行了详细阐述,而这一新发现不仅有助于监测亨廷顿病的进展情况,也有助于开发抑制有害蛋白的新药。 亨廷顿病是一种致命的遗传神经疾病。患者由于基因突变导致机体细胞错误地制造一种名为“变异亨廷顿蛋白”的有害物质,这些异常蛋白会损坏部分脑细胞,导致患者神经系统逐渐退化,致使身体出现不可控制的抽搐,并能发展成痴呆,最后导致死亡。 发表论文的研究小组由英国伦敦大学学院、伦敦国王学院以及诺华生物医学研究所的研究人员组成。他们使用一种新的超敏测试技术测量亨廷顿病患者发病不同阶段体内变异亨廷顿蛋白的水平。这种测试技术名为时间分辨荧光共振能量转移(TR-FRET),可对变异亨廷顿蛋白进行极为精确的测量。研究人员发现,在亨廷顿病患者表现出相关症状之前,其体内变异亨廷顿蛋白即开始逐渐积累,而正常的亨廷顿蛋白含量则在整个发病过程中保持不变。核磁共振扫描显示,亨廷顿病患者脑萎缩的速度明显高于常人。而令研究人员惊讶的是,变异亨廷顿蛋白在白血细胞中的数量与脑萎缩的速率极为一致。这一发现表明,未来或许可以利用血检来预测神经退行性疾病患者脑萎缩的速度。 “使用TR-FRET技术测量变异蛋白水平,是治疗亨廷顿病的一个十分有用的新手段,”研究小组的领导者,伦敦大学学院的萨拉·塔布利兹教授说,“如今获得患者的血液样本很方便,这使得我们能够准确地研究变异亨廷顿蛋白的毒性。而变异亨廷顿蛋白水平与脑萎缩间的关联,使我们对亨廷顿病患者大脑退化的过程有了更新的了解。” 塔布利兹教授指出,TR-FRET技术对于未来基因沉默药物的临床试验也十分有用。她表示,基因沉默药物可用来抑制大脑中有毒蛋白的产生,十分有前途,但也极有可能产生副作用,因此了解药物降低变异亨廷顿蛋白水平的过程十分重要。而TR-FRET技术则提供了这样的手段,这对于新药的开发十分有利。(记者刘海英) 《科技日报》(2012-09-22 二版)
[font=宋体][font=宋体]真核蛋白表达系统是一种广泛应用的蛋白表达方式,通常利用酵母、昆虫或哺乳动物细胞作为宿主。这种表达系统所生成的蛋白与目标[/font][font=Calibri]DNA[/font][font=宋体]具有极高的相似性,能诱导高效蛋白表达。那么,在实施真核蛋白表达时,有哪些关键的纯化步骤呢?接下来,我们将详细解析这一过程。[/font][/font][font=宋体] [/font][font=宋体]首先,我们要明确真核蛋白表达的纯化步骤是至关重要的环节。这些步骤不仅关系到最终产品的纯度和产量,还直接影响其生物活性和应用价值。因此,选择合适的纯化方法对于整个实验的成功至关重要。[/font][font=宋体] [/font][b][font=宋体]真核蛋白表达及纯化步骤主要有以下几个方面:[/font][/b][font=宋体] [/font][font=宋体][font=Calibri]1[/font][font=宋体]、重组质粒构建:将目的基因克隆进表达载体,常见的方法包括限制性切酶切割,基因合成等,根据连接酶说明,进行线性载体和目的基因片段的酶联,最后对质粒测序做好验证;[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2[/font][font=宋体]、蛋白诱导表达:普适条件下查看蛋白是否表达,若不表达,更换载体,表达菌株等方法查看是否表达,如果表达,继续实验;[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3[/font][font=宋体]、蛋白表达部位分析:分析蛋白是可溶性还是不溶性的表达,即在超声后上清表达还是沉淀表达;是否与你的目标蛋白表达部位相同,相同进行后续蛋白表达条件优化;[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4[/font][font=宋体]、蛋白表达优化:优化诱导[/font][font=Calibri]IPTG[/font][font=宋体]浓度、诱导温度,进行放大培养;[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]5[/font][font=宋体]、蛋白纯化:根据目标蛋白的性质进行样本处理,然后进行亲和纯化,获取目的蛋白。[/font][/font][font=宋体] [/font][font=宋体][b]真核表达系统的选择与应用[/b][/font][font=宋体]酵母蛋白表达系统[/font][font=宋体]酵母真核蛋白表达系统有甲醇酵母表达系统,酿酒酵母表达系统,裂殖酵母表达系统以及克鲁维酸酵母表达系统等,其中最早应用于基因工程的酵母是酿酒酵母,但现在运用最广泛的酵母表达系统还是甲醇酵母表达系统中的毕赤酵母真核蛋白表达系统。[/font][font=宋体] [/font][font=宋体]哺乳动物细胞表达系统[/font][font=宋体][font=宋体]哺乳动物细胞表达系统是真核表达系统中唯一可以表达复杂蛋白的系统,它能够指导真核表达蛋白进行正确折叠,提供复杂的[/font][font=Calibri]N[/font][font=宋体]型糖基化和准确的[/font][font=Calibri]O[/font][font=宋体]型糖基化等多种翻译后加工功能,所以它和昆虫酵母系统比较更具有发展潜力,哺乳动物细胞真核表达的蛋白与天然真核表达蛋白的结构、糖基化类型和方式几乎相同且能正确组装成多亚基蛋白[/font][font=Calibri],[/font][font=宋体]但成本较高也一定程度上减缓了它的发展速度。哺乳动物细胞表达系统主要是通过改造宿主细胞来提高外源蛋白的表达效率,常用的宿主细胞有[/font][font=Calibri]CHO[/font][font=宋体]、[/font][font=Calibri]COS[/font][font=宋体]、[/font][font=Calibri]BHK[/font][font=宋体]、[/font][font=Calibri]SP2 /0N[/font][font=宋体]等,哺乳动物转染方法[/font][font=Calibri]*[/font][font=宋体]有脂质体转染法,电穿孔法以及病毒转染等。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州[url=https://cn.sinobiological.com/resource/protein-review/protein-purification-techniques][b]蛋白纯化技术[/b][/url]:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/protein-purification-techniques[/font][/font][font=Calibri] [/font]
酒精废水生产单细胞蛋白技...
1、雷公藤红素抑制CML细胞增殖作者首先进行了网络药理学分析,以评估在治疗CML方面最有效的天然产物。通过对3882种天然产物进行了网络药理学分析,发现从传统中药“雷公藤”(Tripterygium wilfordii)根皮中提取的五环三萜雷公藤红素在抑制CML方面排名第一。为了验证网络药理学筛选的可靠性,作者在CML细胞中进行细胞活力测定。选择18β-甘草次酸作为阴参,因为它与雷公藤红素的结构最相似,但在3882 种天然产物中预测得分不高,选择17-AAG(HSP90抑制剂,已有文章报道HSP90是雷公藤红素的靶点)和TKI 药物伊马替尼作为阳参。结果表明雷公藤红素、17-AAG和伊马替尼均能有效抑制CML细胞增殖,而18β-甘草次酸几乎不影响细胞生长。作者进一步开展细胞实验,发现雷公藤红素对K562和K562T315I细胞表现出抗增殖活性,诱导细胞凋亡。尽管对雷公藤红素的研究很深入,但尚未系统地鉴定出雷公藤红素在CML中的直接蛋白质靶点,尤其是在耐药性CML细胞中 雷公藤红素抑制CML细胞增殖2、雷公藤红素处理后 K562T315I 细胞的转录组和蛋白质组学分析接着,作者通过RNA 测序发现富集的通路包括铁死亡、蛋白水解调节、响应p53介导的DNA损伤等。作者还进行了蛋白质组学分析雷公藤红素对K562T315I细胞中蛋白质表达水平的调节,下调蛋白主要富集于DNA和RNA代谢途径以及 DNA损伤反应,以及蛋白质加工途径。MCODE分析发现“对DNA损伤刺激的反应”和“对未折叠蛋白的反应”分别是最具特征性的途径。雷公藤红素与其已知靶标HSP90的相互作用可能是“对未折叠蛋白的反应”上调的关键贡献事件。而目前尚未有报道称雷公藤红素的直接蛋白质靶标与“对DNA损伤刺激的反应”途径有关( 雷公藤红素处理后 K562T315I 细胞的定量蛋白质组学分析3、雷公藤红素处理后 K562T315I 细胞的CETSA-MS分析作者接着检测了K562T315I细胞中celastrol处理后可溶性蛋白质水平的变化,在雷公藤红素处理后鉴定了178种差异溶解蛋白质,主要位于DNA中心区域,包括细胞核和线粒体,更具体地说是在DNA损伤位点。此外,“分子伴侣复合物”中溶解度降低,这可能是由于雷公藤红素和HSP90之间的互作所致 雷公藤红素处理后 K562T315I细胞的CETSA-MS分析4、雷公藤红素诱导 K562T315I 细胞DNA损伤对 K562T315I细胞经雷公藤红素处理后总蛋白和可溶性蛋白水平变化的系统分析表明,雷公藤红素主要诱导K562T315I细胞中的DNA损伤和未折叠蛋白反应。因此,作者进行了实验来验证这些观察结果。结果显示雷公藤红素显著诱导γ-H2AX(DNA损伤的常见标志物)的表达,并降低DNA损伤修复相关蛋白FANCD2水平,彗星试验进一步证实了雷公藤红素促进的DNA损伤( 雷公藤红素诱导 K562T315I细胞DNA损伤5、雷公藤红素在K562T315I细胞中的靶点鉴定然后,作者在细胞裂解物中开展质谱耦合等温剂量反应-细胞热位移分析(MS-ITDR-CETSA)实验,以确定雷公藤红素的直接蛋白质靶标,特别是那些参与DNA损伤反应的蛋白质靶标。在检测到的3393种蛋白质中,有12种蛋白质表现出热稳定性的显著变化,代表了最有潜力且可信度高的靶标蛋白质。值得注意的是,雷公藤红素的已知靶标HSP90 (HSP90AA1和HSP90AB1) 的热稳定性仅表现出很小的变化,并且没有超过阈值。对这12个潜在靶标和定量蛋白质组学以及CETSA-MS分析的差异蛋白进行PPI分析,发现 YY1均为最紧密相关的蛋白质。因此,YY1与所有这些DEP/DSP的关联节点数量最多,并且可能是与DNA损伤相关的最重要的靶标。现有研究表明,YY1作为转录因子,可以调节参与DNA修复和细胞存活的各种蛋白质的表达,以响应DNA损伤。此外,HMCES已被确定为通过屏蔽脱碱基位点来保护基因组完整性免受氧化碱基损伤的关键蛋白。因此,作者继续通过蛋白质印迹结合细胞热位移分析(WB-CETSA)验证了celastrol与YY1和HMCES的互作。同样,报道的阳性对照HSP90蛋白也显示出明显的热稳定性增加( 雷公藤红素在K562T315I细胞中的靶点鉴定6、雷公藤红素与YY1和HMCES相互作用的验证为了进一步验证celastrol与YY1和HMCES的直接相互作用,合成了可点击炔烃标签功能化celastrol探针(Cel-P),该探针保留了celastrol对K562T315I细胞的抑制活性。利用该探针开展Pulldown实验发现Cel-P 能够成功地从细胞中拉下HMCES和HSP90蛋白,但由于尚不清楚的原因,在蛋白质印迹膜上的下拉样本中未检测到YY1。随后,表达并纯化重组YY1(rYY1)蛋白,发现随着Cel-P浓度的增加,rYY1的标记以剂量依赖性方式增加 雷公藤红素与YY1和HMCES相互作用的验证7、雷公藤红素通过靶向YY1和HMCES诱导DNA损伤在验证了celastrol与YY1和HMCES之间的相互作用后,作者继续在K562T315I细胞中敲低 YY1或HMCES。结果显示YY1或HMCES的敲低显著增加了DNA损伤的发生率,同时影响细胞生长,增强细胞对celastrol的敏感性。此外,与HMCES相比,YY1敲低对细胞的影响更为显著,表明YY1发挥着更为重要的作用。对接分析显示,与HMCES相比,celastrol对YY1的亲和力略强,且Celastrol与YY1上的Leu132和Val316形成氢键,与HMCES的Glu127、Arg130和Arg137形成氢键 雷公藤红素通过靶向 YY1 和 HMCES 诱导 DNA 损伤鉴于YY1在雷公藤红素诱导的DNA损伤反应中发挥关键作用,作者对YY1蛋白进行了进一步实验。发现YY1过表达对细胞生长没有显著影响,但减轻了雷公藤红素引起的细胞死亡和DNA损伤,且通过裂解的PARP1和Caspase-3水平发现YY1表达与雷公藤红素诱导的细胞凋亡呈负相关。使用双荧光素酶报告基因发现雷公藤红素显著抑制了YY1的转录活性,BLI结合试验发现celastrol 可以与 rYY1 结合(图8)。图8 YY1在雷公藤红素诱导的 K562T315I细胞DNA损伤和细胞死亡中起关键作用总结研究使用多组学方法对雷公藤红素的作用机理进行了系统研究,利用蛋白质组范围的无标记靶标反卷积方法MS-CETSA来识别雷公藤红素的蛋白质靶标。研究不仅验证了雷公藤红素通过靶向HSP90来诱导未折叠蛋白反应,而且还发现它通过直接靶向耐药 K562T315ICML 细胞中的YY1和HMCES来诱导DNA损伤(图9)。研究有助于更好地理解雷公藤红素的多方面机制。研究提供了一种有效的系统药理学工作流程范例,该范例集成了网络药理学分析、蛋白质丰度和溶解度测量以及 MS-CETSA,以揭示任何天然产物或活性化合物的作用机理。
俄亥俄州哥伦布市一项新的研究表明,成熟脑细胞表面的三种特定蛋白量的增加可促使细胞产生新的生长延伸。该研究探讨了小鼠脑神经细胞上的三个相关的受体蛋白:GPR3,GPR6和GPR12。当研究人员增加这三种蛋白的量后,细胞生长延伸比蛋白水平正常时的神经细胞的生长大三倍,延伸速度比对照细胞快4-8倍。俄亥俄州立大学医学中心的项目主持人Yoshinaga Saeki说,“我们的研究结果显示,这三种蛋白可能是用于治疗中风、脑和脊髓损伤及神经退行性疾病的重要靶点。”该研究刊登在4月6日的《生物化学杂志》(Journal of Biological Chemistry)上。 这些蛋白量的增加与神经细胞cAMP内的一种重要的信号分子的水平的增加有关。这个分子在调控神经细胞生长、分化和生存,以及传输神经冲动的轴突再生中起着关键作用。随着哺乳动物神经细胞的成熟,其细胞内的cAMP水平下降,这可以部分解释为什么成熟神经细胞受损的轴突不能再生。神经外科副教授、俄亥俄州州立dardinger神经肿瘤及神经科学实验室主管Saeki声称,“我们的发现为cAMP在轴突生长中起着重要作用这一观点提供了更多证据,并显示出这些受体蛋白可能在调节神经细胞cAMP的产生中起主要作用。” 该研究的第一作者Shigeru Tanaka是Saeki所在实验室的一名博士后研究员。在本项研究中,他与同事从小鼠与大鼠脑组织神经母细胞瘤中取得神经细胞,使之在培养基中生长以了解更多关于这三种蛋白及其调控cAMP生长中的作用。他们向这些细胞中注入三种基因以增加这三种蛋白的含量水平,然后用一种被称为核糖核酸干扰的实验室技术关闭这三种蛋白的产生。上述三个蛋白分子中GPR3在神经细胞中最为丰富,而GPR12刺激神经细胞延伸的作用最强。研究表明,阻断GPR3的产生会大大减慢神经细胞的生长速度,研究者们通过修复GPR3或GPR12的产生扭转了这种效应。三种蛋白质的含量水平高也与较高水平的cAMP有关,同时GPR6和GPR12能增加两倍到三倍的水平。 Saeki说,“总的来说,我们的研究结果显示,这三种蛋白能加快神经细胞的生长即使在抑制分子的存在下也是如此,我们迫切希望能找出可以在临床前中风或脊髓损伤动物模型身上重现此结果的方法。”来源:生物谷
美国北卡罗莱纳大学教堂山分校文理学院的首席化学教授约瑟夫—德西蒙博士领导的研究小组发现,人体中的一种正常的良性蛋白质,如果和纳米粒子相结合,就能瞄准并杀死癌细胞,而无须负载那些携带化疗药物的粒子。此前,研究人员曾认为,纳米粒子只有携带了有毒的化学载体才能达到这样的效果。转铁蛋白是人体血液中数量第四多的蛋白质,近20年来一直被作为肿瘤靶向载体用以递送治癌药物。纳米粒子通常也是无毒的,需要通过负载标准化疗药物来治疗癌症。然而,结合转铁蛋白的“打印”纳米粒子,不仅能识别它们,还能诱导癌细胞死亡。而不与任何纳米粒子结合的自由转铁蛋白,能从拉莫斯癌细胞中获得养料生长,即使在很高浓度下也不会杀死任何拉莫斯癌细胞。然而令人吃惊的是,转铁蛋白附着在纳米粒子表面后,其能有效地筛选标靶,攻击并杀死B细胞淋巴瘤。在许多迅速生长的癌细胞表面,蛋白质受体被过度表达,于是和转铁蛋白配体结合的治疗就能找到并瞄准它们,而结合转铁蛋白的纳米粒子被认为是安全且无毒的。德西蒙实验室发明了一种“打印”技术,能人为造出尺寸精确且形状符合预期的纳米颗粒。他们采用这种技术制作出一种可与人类转铁蛋白相结合的生物相容性纳米粒子,其能安全且精确地识别广谱癌症,除了B细胞淋巴瘤外,还能有效地指向非小型细胞,如肺、卵巢、肝脏和前列腺的癌细胞。研究人员目前正在进一步研究,携带转铁蛋白的纳米粒子如何及为何对于拉莫斯癌细胞是有毒的,而对其他细胞却无毒。化学治疗和放射治疗曾被认为是癌症的最有效疗法,但这些疗法通常会损害健康组织和器官。这一发现将可能发展出一种全新的策略来治疗某种类型的淋巴瘤,而副作用更小。不过,德西蒙承认,该研究也会引起一些人对不可预期后果的担忧,即一个设计好的针对某类癌症的靶向化疗载体是否会偏离目标。
[font=宋体][font=Calibri]E. coli[/font][font=宋体]具有遗传背景清楚、细胞增殖快、表达量高、稳定性好和抗污染能力强等特点,适用于多种属蛋白的表达,尤其对小分子蛋白的生产具有极大的优势,但也存在一些问题,如易形成包涵体和含有内毒素等。义翘神州提供从密码子优化到重组蛋白表达[/font][font=Calibri]/[/font][font=宋体]纯化的一站式服务以及内毒素去除等附加服务,以满足不同的定制需求。我们拥有丰富的[/font][font=Calibri]E. coli [/font][font=宋体]可溶性蛋白表达[/font][font=Calibri]/[/font][font=宋体]纯化及蛋白复性经验,拥有多种[/font][font=Calibri]E. coli[/font][font=宋体]细胞株和表达载体,可为客户提供优质的[url=https://cn.sinobiological.com/services/e-coli-protein-expression-service][b]原核蛋白表达服务[/b][/url]。下面是在原核蛋白表达实验中常遇见的几大问题,为大家一一讲解:详情关注:[/font][font=Calibri]https://cn.sinobiological.com/services/e-coli-protein-expression-service[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]1[/font][font=宋体]、我不知道我的蛋白它有什么特性及其结构?[/font][/font][font=宋体] [/font][font=宋体]首先,你要确定一件事,那就是这几个蛋白质有人研究过没有?还是最新发现的蛋白质?如果没有人研究过,那就得用先测部分氨基酸,然后设计引物克隆了。如果有人研究过,那就好了可以根据软件来预测。[/font][font=宋体] [/font][font=宋体][font=宋体]如有[/font][font=Calibri]swiss[/font][font=宋体]—[/font][font=Calibri]pdb[/font][font=宋体]软件,但这个是要有氨基酸序列,知道基因序列,可以在[/font][font=Calibri]ncbi[/font][font=宋体]上进行[/font][font=Calibri]blastx[/font][font=宋体],得到蛋白。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2[/font][font=宋体]、如何选择蛋白表达宿主菌?[/font][/font][font=宋体] [/font][font=宋体]原核系统和真核细胞偏爱的密码子有不同,因此,在用原核系统表达真核基因的时候,真核基因中的一些密码子对于原核细胞来说可能是稀有密码子,从而导致表达效率和表达水平很低。[/font][font=宋体] [/font][font=宋体]原核表达现象:[/font][font=宋体]一、蛋白不表达[/font][font=宋体]①蛋白为毒蛋白[/font][font=宋体]②序列含有稀有密码子[/font][font=宋体] [/font][font=宋体]二、蛋白表达不理想[/font][font=宋体]①蛋白明显降解[/font][font=宋体]②蛋白表达为包涵体[/font][font=宋体]③二硫键错误折叠[/font][font=宋体]④过高的本底表达[/font][font=宋体] [/font][font=宋体][font=Calibri]3[/font][font=宋体]、质粒测序正确,蛋白无法表达怎么办?[/font][/font][font=宋体] [/font][font=宋体][font=宋体]①分析一下稀有密码子,如果比较多,可以尝试[/font][font=Calibri]rosetta[/font][font=宋体]([/font][font=Calibri]DE3[/font][font=宋体]);[/font][/font][font=宋体][font=宋体]②可能是基因本身的问题。[/font][font=Calibri]RNA3[/font][font=宋体]’的特殊结构可能导致转录出现问题,这种情况可以尝试融合表达,譬如[/font][font=Calibri]pET-32a[/font][font=宋体]。[/font][/font][font=宋体][font=宋体]③也许是表达量太低,也可以试一下[/font][font=Calibri]westernblot[/font][font=宋体],定性的检测一下。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4[/font][font=宋体]、如果[/font][font=Calibri]IPTG[/font][font=宋体]诱导后细胞停止了生长,是不是表示细胞死了?[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]T7RNA[/font][font=宋体]聚合酶非常活跃,[/font][font=Calibri]T7[/font][font=宋体]转录和翻译信号极强,因此,一旦诱导,细胞的主要生理活动都向着目的蛋白表达的方面转化。在通常情况下,细胞将停止生长,形成克隆的能力大大降低,但并未死亡。菌落形成试验可以用来检测表达系统的性能。也有一些例外情况,例如特别的目的基因以及一些极为严紧的载体[/font][font=Calibri]/[/font][font=宋体]宿主菌组合(比如含有[/font][font=Calibri]pLysE[/font][font=宋体]的宿主菌)等,这时在诱导后菌落还是会继续生长。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]5[/font][font=宋体]、如何提高重组蛋白在原核细胞里的表达水平,特别是可溶性表达?[/font][/font][font=宋体] [/font][font=宋体]这个问题是最困扰做原核蛋白表达纯化的人的。比如大肠杆菌表达蛋白本身表达量就大,但是表达的大都是包涵体,想要获得可溶性蛋白,就需要做复性,或是再设计实验时就想办法让其在上清中表达。一般就要通过基因优化,载体宿主优化筛选,表达条件优化,诱导条件优化等等。[/font][font=宋体] [/font][font=宋体]①降低重组蛋白合成的速率[/font][font=宋体]可溶性蛋白的产率取决于蛋白的合成速率,蛋白的折叠速率,以及聚集的速率。高水平表达时,肽链聚集的速率一旦超过折叠速率,就会形成包涵体。因此,降低重组蛋白合成的速率有利于提高重组蛋白的可溶性表达。[/font][font=宋体] [/font][font=宋体]②密码子优化[/font][font=宋体][font=宋体]密码子优化就是根据表达系统对密码子的偏好性进行优化筛选。经过优化的基因序列往往能提高[/font][font=Calibri]mRNA[/font][font=宋体]二级结构的稳定性,有利于新生肽段的正确折叠,提高外源活性蛋白的表达。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]③表达温度的选择大肠杆菌的最适生长温度在[/font][font=Calibri]37[/font][font=宋体]~[/font][font=Calibri]39[/font][font=宋体]℃之间,但此温度下极易生成包涵体蛋白,降低可溶性蛋白的表达,而低温培养条件下表达外源蛋白能有效地增加可溶蛋白的比例。[/font][/font][font=宋体] [/font][font=宋体]④诱导条件优化[/font][font=宋体]摇瓶培养时,应选用低菌体浓度诱导,因为在低菌浓度下菌体处于对数生长期,生长活跃,有利于表达可溶性蛋白。然而,如果能保证合理的补料与充分的通气,在较高菌浓度下诱导也同样可能获得可溶蛋白的高效表达。在某些情况下,诱导剂的流加能显著提高可溶蛋白的表达水平。[/font][font=宋体] [/font][font=宋体] [/font][font=宋体][font=Calibri]6[/font][font=宋体]、义翘神州提供标签去除服务吗?[/font][/font][font=宋体] [/font][font=宋体]是的。我们构建载体时可以在标签蛋白和目的蛋白之间加上蛋白酶的酶切位点,这样纯化后就可以利用蛋白酶去除标签,得到完整的目的蛋白。蛋白酶的切割效率受目的蛋白的影响,具体由实验结果而定。[/font][font=Calibri] [/font]
位于HIV病毒表面的糖蛋白gp120可让人体B免疫细胞失去活性,这是《自然—免疫学》Nature Immunology上一项研究给出的结论。这项发现解释了为什么HIV病人抗体反应比较弱。人们已经发现HIV能够引起CD4+辅助性T免疫细胞不断丧失,导致免疫系统功能受损、其他感染更容易乘机而入。但人们一直没有弄清为何在HIV感染早期,T细胞数量还处于相对正常时,人体就不产生有力的抗体反应了。Claudia Cicala与同事发现,HIV的gp120蛋白可通过宿主细胞表面受体α4β7直接作用于B细胞。通过α4β7将信号传递给B细胞,诱发一种免疫反应抑制分子和一种抑制受体的表达。这种由gp120引发的反应大大影响了B细胞活性并降低其产生病毒抗体的能力。
据美国物理学家组织网报道,北卡罗莱纳大学科学家的一项最新发现显示,细胞感染人类疱疹病毒(EBV)后,会产生小泡或被称为外体的液囊,从而改变细胞中所含的蛋白质和RNA(核糖核酸)。这种变质的外体一旦进入健康细胞,就能转变细胞的良性生长方式,使之变成不可控的致癌生长。这一发现刊登于美国《国家科学院院刊》网络版。EBV可能是世界上最成功的病毒,它无法被免疫系统彻底清除,几乎每个人终生都被它感染。它们不断进入唾液,在这里进行有效地传播。感染这种病毒很少致病,然而在几种主要的癌症中都发现了它的踪迹,包括淋巴瘤和鼻咽癌,它的蛋白质劫持了细胞生长调控机制,引发不可控的细胞生长,从而导致癌变。研究认为一种名为潜伏膜蛋白质1的蛋白质是EBV的致癌基因。通过外体,它们被传递给未受感染的细胞。研究人员还指出,EBV也彻底改变了外体的内含物,在细胞之间传递能激活癌症的蛋白质,这是值得注意的地方。这些发现表明,通过这种方式,病毒感染细胞能广泛影响并潜在控制全身其他细胞,引发它们的不可测生长。免疫系统不断地监视着外来病毒蛋白质,然而经外体携带的这些蛋白质可以不向免疫系统“报告”感染,并刺激癌细胞生长,由此容许了一种不可测的生长。该研究还显示,细胞能产生血管,这一被称为血管新生的过程很容易接受变质外体并引发潜在生长。北卡罗莱纳大学莱恩伯格综合癌症研究中心微生物与免疫学教授南希·瑞玻-特拉玻说:“外体就像特洛伊木马,EBV通过木马甚至能控制那些还没有感染的细胞。但重要的是,外体的产生可能为我们提供了一种新的治疗标靶,封锁它们就能控制癌症蔓延。”论文第一作者、瑞玻-特拉玻实验室博士后戴维·麦克表示,下一步研究是测定哪些蛋白质被选中进入外体,病毒如何控制了这些蛋白质,以及怎样才能遏制这一过程。
转铁蛋白与纳米粒子结合就可瞄准并杀死拉莫斯癌细胞,而无需负载其他化疗药物,此项发现将有望发展出癌症靶向治疗的新策略。 相关研究成果发表在本周的《美国化学协会杂志》上。 美国北卡罗莱纳大学教堂山分校文理学院的首席化学教授约瑟夫德西蒙博士领导的研究小组发现,人体中的一种正常的良性蛋白质,如果和纳米粒子相结合,就能瞄准并杀死癌细胞,而无须负载那些携带化疗药物的粒子。此前,研究人员曾认为,纳米粒子只有携带了有毒的化学载体才能达到这样的效果。 转铁蛋白是人体血液中数量第四多的蛋白质,近20年来一直被作为肿瘤靶向载体用以递送治癌药物。纳米粒子通常也是无毒的,需要通过负载标准化疗药物来治疗癌症。然而,结合转铁蛋白的“打印”纳米粒子,不仅能识别它们,还能诱导癌细胞死亡。而不与任何纳米粒子结合的自由转铁蛋白,能从拉莫斯癌细胞中获得养料生长,即使在很高浓度下也不会杀死任何拉莫斯癌细胞。 然而令人吃惊的是,转铁蛋白附着在纳米粒子表面后,其能有效地筛选标靶,攻击并杀死B细胞淋巴瘤。在许多迅速生长的癌细胞表面,蛋白质受体被过度表达,于是和转铁蛋白配体结合的治疗就能找到并瞄准它们,而结合转铁蛋白的纳米粒子被认为是安全且无毒的。 德西蒙实验室发明了一种“打印”技术,能人为造出尺寸精确且形状符合预期的纳米颗粒。他们采用这种技术制作出一种可与人类转铁蛋白相结合的生物相容性纳米粒子,其能安全且精确地识别广谱癌症,除了B细胞淋巴瘤外,还能有效地指向非小型细胞,如肺、卵巢、肝脏和前列腺的癌细胞。 研究人员目前正在进一步研究,携带转铁蛋白的纳米粒子如何及为何对于拉莫斯癌细胞是有毒的,而对其他细胞却无毒。 化学治疗和放射治疗曾被认为是癌症的最有效疗法,但这些疗法通常会损害健康组织和器官。这一发现将可能发展出一种全新的策略来治疗某种类型的淋巴瘤,而副作用更小。 不过,德西蒙承认,该研究也会引起一些人对不可预期后果的担忧,即一个设计好的针对某类癌症的靶向化疗载体是否会偏离目标。(科技日报)
将在药物开发进程中发挥重要作用2011年03月26日 来源: 科技日报 作者: 常丽君 本报讯 据每日科学网近日报道,美国范德堡大学研究人员开发出一种新型激光技术,可检测细胞膜上的蛋白质和其它多种生物分子之间的相互反应。这种检测将在药物开发进程中发挥重要作用。 人类细胞中约有7000种蛋白质,其中30%在细胞膜上,控制细胞分子运作机制的信号有60%—70%由这些膜蛋白产生,因此当前市场上约一半的药物都是瞄准细胞膜蛋白。但因为膜蛋白很难提纯,科学家在研究它们的结构时面临很多困难。现有的检测膜手段大多是将膜蛋白从其所处环境中分离,或用不同方式如荧光标签加以修改,以分析它们的活性。这些方法不仅昂贵耗时,还可能会影响目标膜蛋白的功能。 范德堡大学化学生物研究院化学教授达里尔·波恩霍普领导的研究小组和斯克里普斯研究院合作,开发了一种名为“后向散射干涉仪”(BSI)的新型激光技术,能精确检测出膜蛋白和自然界中各种分子之间的结合力。 BSI操作起来很简单,只要把两种分子混合装入一个充满液体的显微镜小盒中,用一束类似于条形码扫描仪的红色激光照射,就能测出它们之间的结合力。小盒的几何形状调整合适后,激光就会产生干涉图案,而这种干涉图案对分子之间的反应非常敏感。如果分子开始互相作用,图案就开始变换。 为了检验BSI的准确性,研究人员制造了一种含有GM1小蛋白质的合成膜,霍乱毒素要进入细胞,主要结合对象就是这种小蛋白质。他们把霍乱毒素B和这些膜混合,检测出的结合力结果与用其他方法所得到的结果一致。为了进一步确认,他们还用了一种和胸腺癌相关的天然分离膜和3种分别与疼痛、发炎和神经传导素GABA(用于放松、睡眠和调节紧张)相关的蛋白质膜进行检验,把包含这些蛋白质的膜和对应结合分子相混合,用BSI技术测得的值也和用其他方法得到的结果一样。 此外,该技术进入商业化也前景广阔,范德堡大学对新型激光检测技术已申请了专利,并已获得3项批准。他们还专门成立了一家分子传感公司对新技术进行独家开发。
我正在做细胞色素C 胰蛋白酶酶解后[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]测序,我想请教一下酶解的条件。酶的用量,时间,温度,pH,还有缓冲溶剂的配置
Scientists Ratchet Up Understanding of Cellular Protein Factory科学家对于分子蛋白工厂的理解ScienceDaily (Dec. 1, 2010) — Theoretical biologists at Los Alamos National Laboratory have used a New Mexico supercomputer to aid an international research team in untangling another mystery related to ribosomes -- those enigmatic jumbles of molecules that are the protein factories of living cells. The research, published December 2 in the journal Nature, could aid in development of new antibiotics used to fight multidrug resistant superbugs such as MRSA (methicillin-resistant Staphylococcus aureus infections) found in many U.S. hospitals. The work may also be important for combating engineered strains of anthrax and plague.科学日报(2010年12月1日) - 在美国洛斯阿拉莫斯国家实验室的理论生物学家已经使用了新墨西哥州的超级计算机,以帮助一个国际研究小组解开有关核糖体另一个谜- 活细胞的蛋白质工厂分子神秘动力 。这项研究发表在12月2日的自然杂志上,可以有助于研发药针对美国许多医院中的对抗生素抗药的葡萄球菌(耐甲氧西林金黄色葡萄球菌感染)。这项工作也可用于防治炭疽工程菌和鼠疫。In the context of synthetic biology, understanding the ribosome could be key to developing nanofactories that produce designer biomolecules and polymers.在生物合成方面,了解核糖体是关键,发展纳米工厂来生产所设计的生物分子和聚合物。In the paper, "Head swivel on the ribosome facilitates translocation via intra-subunit tRNA hybrid sites," Los Alamos National Laboratory researchers Karissa Sanbonmatsu and Paul Whitford and José N. Onuchic at the University of California-San Diego join Christian Spahn, Andreas Ratje, and others from the Institute for Medical Physics and Biophysics, Berlin, Germany, to describe for the first time how a complicated swivel movement within a bacterial ribosome accommodates synthesis of proteins.在论文中,“头部旋转有利于通过内部核糖体亚基的tRNA移位”, 加州大学圣迭戈分校洛斯阿拉莫斯国家实验室的研究人员卡Karissa Sanbonmatsu 、 Paul Whitford 和 José N. Onuchic,其他来自柏林,德国医学物理学和生物物理学研究所,来形容首次如何在细菌核糖体进行复杂旋转运动来合成蛋白质。Ribosomes are composed of long chemical chains, called ribonucleic acids (RNA), and proteins. Each ribosome has two interlocked subunits, one large and one small, which behave as a single molecular machine. Because of its makeup, each ribosome resembles a tangle of threads or a handful of rubber bands tossed together. Despite the ribosome's outwardly disjointed appearance, researchers have found that the two subunits ratchet, un-ratchet, and swivel during protein synthesis to allow introduction of helper chemicals called transfer RNAs (tRNAs) into its folds to manufacture new chains of protein molecules. The proteins are used to create new cells or perform necessary functions within the host cell or organism.核糖体是由称为(RNA)的核糖核酸和蛋白质长化学链构成。每个亚基核糖体有两个相关的亚单位,一大一小,这表现为一个单一的分子机器。由于其构成,每个核糖体类似于一个线程缠结或一把橡皮筋捆在一起。尽管核糖体的表面上出现脱节,研究人员发现,这两个亚基有助于转运RNA(tRNA基因)来制造新的蛋白质分子链。这些蛋白质被用来制造新的细胞或在宿主细胞内或者生物体中执行必要的功能。Ribosomes build proteins by linking chemical segments fashioned from instructions delivered via messenger RNA, which is DNA's molecular cousin. Each segment, or amino acid, corresponds to a trio of bases in the message that, in turn, complement trios encoded in transfer RNA. Each base in the trio corresponds to a single chemical complement found on the RNA. In order for protein synthesis to occur, the tRNA must bind to the ribosome at two distinct sites -- one to decode the information and another to link the new amino acid to the emerging protein.核糖体从通过信使RNA将化学片段链接来制造蛋白,这是DNA的分子交付。每一部分,或氨基酸,对应于信使中的三个碱基,反过来,转运RNA编码的三个互补碱基。三人中的每个碱基都对应RNA上的一个化学成分。为了使蛋白质的合成,tRNA必须在两个不同位点结合到核糖体 - 一个解码信息,另一个连接新的氨基酸去合成新的蛋白质。After each amino acid is added, the ribosome must crawl along the message to create additions. Exactly how this crawling occurs has been a mystery for several decades. Researchers have suspected that ratcheting motions of the two ribosomal subunits are key to allowing RNA and associated catalysts into the complex structure of the ribosome so the RNA and ribosome can couple at the crucial sites to create proteins. In the Nature paper, the researchers discovered that the majority of crawling (movement along messenger RNA) occurs during a new kind motion, "head swivel," rather than ratcheting.每个氨基酸添
1月2日,佛山奥素博新科技有限公司(以下简称奥素科技)宣布完成近亿元A轮融资。本轮融资由鲁信创投领投,老股东启明创投、线性资本、同创伟业等持续加码,凯乘资本(WinX Capital)担任财务顾问。本轮融资后,奥素科技将进一步加速在单细胞蛋白组学领域的商业化推广,提供差异化的产品和服务,填补实验室样本预处理、功能发现及验证等需求的空白,力争将中国制造的先进生命科学仪器推向全球市场。奥素科技成立于2021年,具有全球领先的有源数字微流控液滴操控平台,在两年多时间内已连续获得四轮融资,股东包括诸多顶级VC及知名产业投资人。公司推出的第一款商业化产品Boxmini? SCP,是全球首款全流程微流控片上单细胞蛋白组学样本前处理工具,高效协助用户实现高通量、快速、精确的微量样本控制,一站式完成复杂的单细胞蛋白质样本前处理工作,且对无标记和TMT标记处理方案均可适配,产品推出后备受市场关注。[img=图片]https://img1.17img.cn/17img/images/202401/uepic/ea05f042-359a-4e3c-a6db-2edf2fa796f0.jpg[/img]对于本次融资,[b]奥素科技创始人兼CEO马汉彬博士[/b]表示:“将消费电子半导体技术引入到生命科学领域,奥素团队已经完成了0到1的积累:特别是在单细胞蛋白质组学样本前处理应用场景,我们通过有源数字微流控微芯片上纳升样本精准操控及全流程集成能力,获得了海内外多位头部PI的认可并产生了对整个领域有促进意义的实验结果;在单细胞多组学、微生物及合成生物学等其他领域,奥素也将与不同的下游伙伴携手前行,加速新产品的开发及商业化落地。我们将在新老股东的支持下,利用产品技术优势,迅速开拓海内外市场,以单细胞蛋白质组学产品为突破点,通过开放式数字微流控共享平台打造半导体技术的生物芯片生态,让生命科学实验室及医疗检验自动化快速迈入消费电子时代。”此前,在仪器信息网[url=https://www.instrument.com.cn/webinar/meetings/icca2023][font=arial][color=#000000]第六届细胞分析网络大会(iCCA2023)的【单细胞分析技术】专题会场中,[/color][/font][/url][url=https://www.instrument.com.cn/webinar/meetings/icca2023]马汉彬[/url]研究员分享《[url=https://www.instrument.com.cn/webinar/meetings/icca2023][font=arial][color=#c00000] [/color][/font][color=#c00000]基于有源数字微流控的单细胞分选和操控系统[/color][/url]》的主题报告。[url=https://www.instrument.com.cn/webinar/meetings/icca2023][i](详情点击)[/i][/url][align=center][url=https://www.instrument.com.cn/webinar/meetings/icca2023][img=,200,200]https://img1.17img.cn/17img/images/202308/uepic/34781884-5a9f-4e5a-b5df-03783a77c663.jpg[/img][/url][/align][align=center][url=https://www.instrument.com.cn/webinar/meetings/icca2023]马汉彬 中国科学院苏州生物医学工程技术研究所 研究员[/url][/align]马汉彬研究员课题组也在2023年成功研发出了一套基于大面积薄膜晶体管开关阵列的有源数字微流控平台,在Analytical Chemistry发表并被选为当期的封面论文。[url=https://www.instrument.com.cn/news/20230817/680090.shtml][i](详情点击)[/i][/url][align=center][img=7872d6238fc05517bb5f145142a71dee_7b4ffe77-43f0-48a2-886a-b31b248d32ca.jpg,350,465]https://img1.17img.cn/17img/images/202401/uepic/e362de5d-8580-4bc7-baf3-7272a8a7d52c.jpg[/img][/align]本轮领投方,[b]鲁信创投副总经理邱方[/b]表示:“鲁信创投作为国有控股的专业创投机构,一贯秉持以创业投资形式,支持我国自主的研究平台、仪器设备成果应用转化,将实现我国高水平科技自立自强的任务放在首位。奥素科技掌握有源数字微流控的核心底层技术,有潜力将实验室自动化推进到一个全新的局面,形成新的研究平台。公司推出的单细胞蛋白组学产品,为单细胞多组学等前沿研究提供先进工具,在包括鲁信已投企业在内的下游客户中引起强烈关注,体现出国产科学仪器的高水平自立自强,即将迎来新的局面。鲁信创投将支持奥素科技,打好科学仪器设备国产化攻坚战。”[b]启明创投合伙人陈侃[/b]表示:“启明创投作为上轮领投方,已连续两轮增资奥素科技。公司凭借强大的研发能力和优秀的执行力,快速的推出了单细胞领域的尖刀产品,面向一片蓝海市场。我们对公司未来充满信心,继续助力公司海外市场的商业化,期待奥素科技将“中国智造”先进科学仪器推向世界。”[b]线性资本董事总经理郑灿[/b]表示:“线性资本作为天使轮领投方,坚定认为投资要找到正确的人。我们亲眼见证了马汉彬博士从一名科研工作者向现代企业家的转变。马汉彬博士的为人、科学素养、前沿视野和企业家精神令我们印象深刻。在他带领下,公司首先推出了具有划时代意义的单细胞蛋白质组学解决方案,为全球蛋白组学领域研究再填一把火。我们本轮继续增持,推动奥素科技向先进科学仪器标杆企业迈进。”[b]同创伟业北京医药基金合伙人郗砚彬[/b]表示:“我们始终认为,奥素科技的数字微流控芯片系统,有望成为下一代生命科学微反应器的关键载体,持续为科学研究、医药工业等提供创新解决方案。公司的单细胞蛋白组学产品,将蛋白组学研究推进到了切实可行的单细胞颗粒度,使客户能够不再受工具所限,以全新的角度验证所知和探索未知。我们本轮继续增持,期待奥素科技能够让先进技术在应用层面全面开花。”[b]凯乘资本创始合伙人邹国文[/b]表示:“凯乘资本很荣幸连续第三轮担任奥素科技融资的财务顾问,见证了奥素从初创、一路飞速发展及商业化;作为数字微流控行业头部企业,奥素能够穿越市场周期,在不到三年的时间连续获得四轮融资,充分体现了资本端对公司的高度认可。期待奥素在下游领域的进一步拓展,成为世界领先的生命科学工具企业。”[b]关于鲁信创投:[/b]鲁信创投是山东省鲁信投资控股集团有限公司控股的省内最大、国内具有重要影响力的专业创投机构,是国内资本市场首家上市的创投机构(股票代码:600783.SH)。成立20余年以来,管理运作各类基金已达40余只,基金规模约200亿元,覆盖医疗健康、军民融合、先进制造、电子信息、新能源、新材料等细分产业,境内外上市公司40余家,在医疗健康领域先后投资了思路迪、硅基仿生、中科新生命、爱博泰克、唯迈医疗、美东汇成、英赛斯、荣昌生物等一批优秀企业。[来源:仪器信息网] 未经授权不得转载[align=right][/align]
[font=宋体][font=宋体]重组蛋白([/font][font=Calibri]recombinant protein[/font][font=宋体])是指应用重组 [/font][font=Calibri]DNA [/font][font=宋体]或重组 [/font][font=Calibri]RNA [/font][font=宋体]技术而获得的蛋白质。重组蛋白工程先应用基因克隆或化学合成技术获得目的基因([/font][font=Calibri]gene of interest[/font][font=宋体],[/font][font=Calibri]GOI[/font][font=宋体]),连接到适合的表达载体,导入到特定的宿主细胞,利用宿主细胞的遗传系统,表达出有功能的蛋白质分子。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]重组蛋白的产生是应用了重组[/font][font=Calibri]DNA[/font][font=宋体]或重组[/font][font=Calibri]RNA[/font][font=宋体]的技术从而获得的蛋白质。体外重组蛋白的生产主要包括四大系统:原核蛋白表达,哺乳动物细胞蛋白表达,酵母蛋白表达及昆虫细胞蛋白表达。生产的蛋白在活性和应用方法方面均有所不同。根据自身的下游运用选择合适的蛋白表达系统,提高表达成功率。[/font][/font][font=宋体] [/font][font=宋体]其获得途径可以分为体外方法和体内方法。两种方法的前提都是应用基因重组技术,获得连接有可以翻译成目的蛋白的基因片段的重组载体,之后将其转入可以表达目的蛋白的宿主细胞从而表达特定的重组蛋白分子。[/font][font=宋体] [/font][font=宋体][font=宋体][b]当前重组蛋白的生产主要有四大系统[/b]:原核表达系统:最常用的大肠杆菌蛋白表达,真核表达系统如酵母,哺乳动物细胞蛋白表达(常用的细胞[/font][font=Calibri]CHO[/font][font=宋体],[/font][font=Calibri]HEK293[/font][font=宋体])及、昆虫细胞蛋白表达系统。重组蛋白的产生尚可利用转基因动物的乳腺或者植物产生,产生的重组蛋白作为生物制药的产物,在医学中作用显著。利用基因工程技术,可以使细胞或者动物本身变成“批量生产药物的工厂”。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]以利用转基因动物的乳腺表达重组蛋白为例:其方法是将药用蛋白基因与乳腺蛋白基因的启动子等调控组件重组在一起,通过显微注射等方法,导入哺乳动物(哺乳动物才会泌乳)的受精卵中,然后,将受精卵送入母体内,使其生长发育成转基因动物。转基因动物进入泌乳期后,可以通过分泌的乳汁来生产所需要的蛋白质药品,因而称为动物乳腺生物反应器或乳房生物反应器。科学家已在牛和山羊等动物的乳腺生物反应器中表达出了抗凝血酶、血清白蛋白、生长激素和[/font][font=宋体]α[/font][font=Calibri]-[/font][font=宋体]抗胰蛋白酶等重要的医药产品。[/font][/font][font=宋体]重组蛋白在制药工业上主要是指表达获得的细胞因子、凝血因子或者人工设计的蛋白分子。[/font][font=宋体] [/font][font=宋体][font=宋体]目前,重组蛋白试剂已被广泛应用于生物药、细胞免疫治疗及诊断试剂的研发和生产中。其中重组蛋白药物是生物药物的重要组成成分,常被被广泛应用于医疗领域[/font][font=Calibri],[/font][font=宋体]包括肿瘤治疗、免疫调节、神经保护、结缔组织疾病、肾病治疗等。包括细胞因子类、抗体治疗性疫苗、激素及酶等。[/font][/font][font=宋体] [/font][font=宋体]义翘神州致力于提供[url=https://cn.sinobiological.com/resource/protein-review/protein-production][b]重组蛋白生产[/b][/url]、[url=https://cn.sinobiological.com/resource/protein-review/protein-expression][b]重组蛋白表达[/b][/url]及[url=https://cn.sinobiological.com/resource/protein-review/protein-production-systems][b]重组蛋白系统[/b][/url]详情的咨询与解决方案。为实验中特定的应用选择正确的表达系统是成功的关键所在。在选择表达系统时,蛋白溶解度、功能、纯化速度和产量通常是必须考虑的重要因素。此外,每个表达系统都有其独特的优势和挑战,这一点在选择时也需着重考虑。我们的专业团队将为您提供个性化的建议,以帮助您根据实验需求选择最合适的表达系统。[/font][font=宋体][font=宋体]更多详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/protein-production[/font][/font][font=Calibri] [/font]
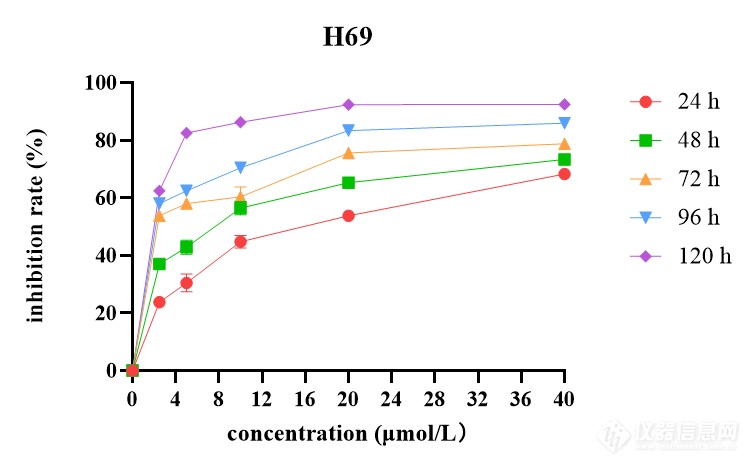
[align=left][size=18px]西达本胺促进[/size][size=18px]S[/size][size=18px]CLC[/size][size=18px]细胞系组蛋白乙酰化[/size][/align][align=left][size=18px] [/size][size=18px] [/size][size=16px]为验证西达本胺是否上调[/size][size=16px]SCLC[/size][size=16px]细胞系的乙酰化水平,我们使用[/size][size=16px]Western blot[/size][size=16px]检测了不同浓度([/size][size=16px]I[/size][size=16px]C10[/size][size=16px]、[/size][size=16px]IC20[/size][size=16px]、[/size][size=16px]IC50[/size][size=16px])西达本胺处理[/size][size=16px]4[/size][size=16px]8[/size][size=16px] [/size][size=16px]h[/size][size=16px]后,[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞系中乙酰化组蛋白[/size][size=16px]H[/size][size=16px]3[/size][size=16px]、[/size][size=16px]H[/size][size=16px]4[/size][size=16px]表达水平,并以组蛋白[/size][size=16px]H[/size][size=16px]3[/size][size=16px]、[/size][size=16px]H[/size][size=16px]4[/size][size=16px]表达水平为对照。结果如图所示。在四种亚型细胞系中,总组蛋白[/size][size=16px]H[/size][size=16px]3[/size][size=16px]、[/size][size=16px]H[/size][size=16px]4[/size][size=16px]表达水平无变化,乙酰化组蛋白[/size][size=16px]H[/size][size=16px]3[/size][size=16px]、[/size][size=16px]H[/size][size=16px]4[/size][size=16px]表达量随加药浓度增大而增多,这证明了西达本胺对[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞系组蛋白乙酰化的促进作用,这种作用呈剂量依赖性。[/size][/align][align=left][size=18px]A[/size][/align][align=left][img]https://ng1.17img.cn/bbsfiles/images/2022/11/202211302350400898_8640_5887180_3.png[/img][/align][align=left][/align][align=left][/align][align=left][/align][align=center][/align][align=left][size=18px] [/size][size=18px]西达本胺通过线粒体凋亡途径诱导[/size][size=18px]S[/size][size=18px]CLC[/size][size=18px]细胞系凋亡[/size][/align][align=left][size=16px]我们的功能实验表明,西达本胺[/size][size=16px]可剂量依赖的[/size][size=16px]促进[/size][size=16px]SCLC[/size][size=16px]细胞[/size][size=16px]系[/size][size=16px]凋亡[/size][size=16px],但其机制尚未明确。[/size][size=16px]依据国内外报道,西达本胺主要通过线粒体凋亡途径诱导细胞凋亡[/size][size=16px]。除此之外,[/size][size=16px]西达本胺[/size][size=16px]能[/size][size=16px]使[/size][size=16px]线粒体[/size][size=16px]DNA[/size][size=16px]双链断裂,发生损伤。[/size][size=16px]为探究其是否通过此途径在[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞系中发挥作用,我们检测了加药[/size][size=16px]4[/size][size=16px]8[/size][size=16px] [/size][size=16px]h[/size][size=16px]后,[/size][size=16px]H69[/size][size=16px]、[/size][size=16px]H[/size][size=16px]446[/size][size=16px]、[/size][size=16px]H[/size][size=16px]526[/size][size=16px]、[/size][size=16px]DMS114[/size][size=16px]细胞中由线粒体介导的[/size][size=16px]C[/size][size=16px]aspase[/size][size=16px]信号通路相关蛋白[/size][size=16px]Bcl-2[/size][size=16px],[/size][size=16px]Bax[/size][size=16px],细胞色素[/size][size=16px]C[/size][size=16px],[/size][size=16px]Ca[/size][size=16px]spase 9[/size][size=16px],[/size][size=16px]c[/size][size=16px]leaved Caspase 9[/size][size=16px],[/size][size=16px]P[/size][size=16px]ARP[/size][size=16px],[/size][size=16px]c[/size][size=16px]leaved [/size][size=16px]PARP[/size][size=16px],[/size][size=16px]Ca[/size][size=16px]spase 3[/size][size=16px],[/size][size=16px]c[/size][size=16px]leaved Caspase 3[/size][size=16px]以及[/size][size=16px]D[/size][size=16px]NA[/size][size=16px]双链断裂标志物[/size][size=16px] [/size][size=16px]γH2AX[/size][size=16px]表达水平。[/size][size=16px]Western blot[/size][size=16px]结果显示,[/size][size=16px]Ca[/size][size=16px]spase 9[/size][size=16px],[/size][size=16px]P[/size][size=16px]ARP[/size][size=16px] [/size][size=16px],[/size][size=16px]Ca[/size][size=16px]spase 3[/size][size=16px]表达水平无明显变化,[/size][size=16px]Bcl-2[/size][size=16px]表达下调,其余蛋白表达均上调。这些结果表明,在[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞中,西达本胺可以通过线粒体凋亡途径诱导细胞凋亡。[/size][/align][align=left][/align][align=left][/align][align=left][/align][align=left][/align][align=left][/align][align=left][/align][align=left][/align][align=left][/align][align=left][size=16px]A[/size][/align][align=left][img]https://ng1.17img.cn/bbsfiles/images/2022/11/202211302350402755_79_5887180_3.png[/img][/align][align=left][size=18px] [/size][size=18px]西达本胺通过抑制[/size][size=18px]C[/size][size=18px]yclin-CDK[/size][size=18px]复合物活性阻滞[/size][size=18px]S[/size][size=18px]CLC[/size][size=18px]细胞系周期[/size][/align][align=left][font='宋体'][size=16px]据文献报道,[/size][/font][size=16px]不同[/size][size=16px]HDACI[/size][size=16px]对不同细胞阻滞时相不一致。为验证西达本胺对[/size][size=16px]SCLC[/size][size=16px]细胞周期的作用,我们检测了[/size][size=16px]经[/size][size=16px]西达本胺[/size][size=16px]处理[/size][size=16px]48[/size][size=16px] [/size][size=16px]h[/size][size=16px]后,[/size][size=16px]H69[/size][size=16px]、[/size][size=16px]H446[/size][size=16px]、[/size][size=16px]H526[/size][size=16px]、[/size][size=16px]DMS114[/size][size=16px]细胞中细胞周期相关蛋白的表达水平,如图所示。[/size][size=16px]在[/size][size=16px]H69[/size][size=16px]、[/size][size=16px]H526[/size][size=16px]、[/size][size=16px]D[/size][size=16px]MS114[/size][size=16px]细胞系中[/size][size=16px]P21[/size][size=16px]、[/size][size=16px]P27[/size][size=16px]表达上调,[/size][size=16px]C[/size][size=16px]yclin A2[/size][size=16px]与[/size][size=16px]C[/size][size=16px]DK[/size][size=16px]2[/size][size=16px]表达下调[/size][size=16px],[/size][size=16px]说明西达本胺阻滞[/size][size=16px]H69[/size][size=16px]、[/size][size=16px]H526[/size][size=16px]、[/size][size=16px]D[/size][size=16px]MS114[/size][size=16px]于[/size][size=16px]S[/size][size=16px]期。在[/size][size=16px]H446[/size][size=16px]细胞系中[/size][size=16px]C[/size][size=16px]yclin E1[/size][size=16px]与[/size][size=16px]C[/size][size=16px]DK2[/size][size=16px]表达下调[/size][size=16px],说明西达本胺阻滞其于[/size][size=16px]G[/size][size=16px]1[/size][size=16px]/S[/size][size=16px]期。[/size][/align][align=left][img]https://ng1.17img.cn/bbsfiles/images/2022/11/202211302350422585_1956_5887180_3.png[/img][size=16px] [/size][img]https://ng1.17img.cn/bbsfiles/images/2022/11/202211302350405804_8826_5887180_3.png[/img][/align][align=left][size=18px]小结[/size][/align][size=16px]1[/size][size=16px].[/size][size=16px]西达本胺可以增强[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞系组蛋白乙酰化水平。[/size][size=16px]2.[/size][size=16px]西达本胺诱导[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞凋亡的机制可能与其激活线粒体介导的[/size][size=16px]caspase[/size][size=16px]凋亡途径有关。[/size][size=16px]3[/size][size=16px].[/size][size=16px]西达本胺可阻滞[/size][size=16px]S[/size][size=16px]CLC[/size][size=16px]细胞周期,可能与其上调细胞周期蛋白激酶抑制剂表达、从而抑制[/size][size=16px]C[/size][size=16px]yclin-CDK[/size][size=16px]复合物活性有关。[/size]
[b][font=宋体]整合蛋白和跨膜蛋白定义:[/font][/b][font=宋体] [/font][font=宋体]整合蛋白和跨膜蛋白是两类重要的蛋白质,它们在细胞分子水平上起着重要的作用。[/font][font=宋体] [/font][font=宋体]整合蛋白,也称为内在蛋白或跨膜蛋白,部分或全部镶嵌在细胞膜中或内外两侧,以非极性氨基酸与脂双分子层的非极性疏水区相互作用而结合在质膜上。它们是生物膜的基本结构成分,许多具重要生理功能的膜蛋白均属整合蛋白,如膜结合的酶类、载体蛋白、通道蛋白、膜受体等。[/font][font=宋体] [/font][font=宋体]跨膜蛋白,是可以跨越细胞膜的蛋白,它在细胞的信号传递系统中担当着重要的角色。跨膜蛋白在结构上可以分为单次跨膜、多次跨膜、多亚基跨膜等,它们具有能够跨越细胞膜的能力。[/font][font=宋体] [/font][b][font=宋体]整合蛋白和跨膜蛋白在位置、结构和功能上存在显著的差异[/font][/b][font=宋体] [/font][font=宋体]①位置:整合蛋白主要存在于细胞质内,细胞核或其他非细胞膜结构中,它们容易在细胞中自由移动。而跨膜蛋白则嵌入细胞膜中,一部分位于细胞膜的胞外侧,另一部分位于细胞膜的胞内侧,形成了一个穿过细胞膜的通道。[/font][font=宋体][font=宋体]②结构:整合蛋白的结构通常由两个独立的部分组成,一个是靠近细胞膜的膜结合区域([/font][font=Calibri]TM[/font][font=宋体]),另一个是靠近细胞骨架的非膜结合区域([/font][font=Calibri]N-TM[/font][font=宋体])。当接受到外界的信号时,整合蛋白的[/font][font=Calibri]TM[/font][font=宋体]区域会被激活,把来自外界的信号转化为细胞内可以识别的信号,直接参与细胞信号传导系统中。[/font][/font][font=宋体]③功能:整合蛋白主要是用来从外界传达信号到细胞内,充当细胞与外界信号的桥梁。而跨膜蛋白则在细胞的信号传递系统中担当着重要的角色。[/font][font=宋体]总的来说,整合蛋白和跨膜蛋白在位置、结构和功能上存在显著的差异,这些差异使得它们在生物体中扮演着不同的角色。[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/resource/protein-review/transmembrane-proteins][b]跨膜蛋白表达与制备服务[/b][/url],制备流程图:基因合成[/font][font=宋体]→载体构建→细胞转化[/font][font=Calibri]/[/font][font=宋体]转染→蛋白表达→细胞收集→细胞破碎→膜脂提取→膜脂增溶→蛋白纯化→质量检测,同时义翘拥有[/font][/font][b][font=宋体]三大跨膜蛋白制备平台[/font][/b][font=宋体],可以为客户提供全面的多次跨膜蛋白产品和服务。同时,为基础研究和药物研发提供更加优质的原材料。[/font][font=宋体] [/font][b][font=宋体][font=Calibri]VLP[/font][font=宋体]技术平台[/font][/font][/b][font=宋体][font=宋体]正确折叠的膜蛋白在细胞膜上表达,类病毒颗粒[/font][font=Calibri]VLP[/font][font=宋体]通过出芽的方式包裹上携带有靶标蛋白的细胞膜,形成包膜的[/font][font=Calibri]VLP[/font][font=宋体]。它是由病毒的衣壳蛋白通过自组装而形成的纳米级颗粒(直径约[/font][font=Calibri]100[/font][font=宋体]~[/font][font=Calibri]300[/font][font=宋体]纳米),不含病毒核酸,不能进行自主复制,生产操作过程中较为安全。产生的[/font][font=Calibri]VLP[/font][font=宋体]蛋白可直接像可溶蛋白一样进行包被进行[/font][font=Calibri]ELISA[/font][font=宋体]检测。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州已成功开发[/font][font=Calibri]VLP[/font][font=宋体]技术平台,它可以将完整天然构象的膜蛋白展示在类病毒颗粒表面,这种方法不仅可以保留膜蛋白的完整结构,同时也能够真实地模拟其在细胞膜上的位置和构象。[/font][/font][font=宋体] [/font][b][font=宋体]去垢剂技术平台[/font][/b][font=宋体][font=宋体]由于存在疏水结构域,跨膜蛋白与膜的结合非常紧密,需要用去垢剂([/font][font=Calibri]detergent[/font][font=宋体])才能从膜上洗涤下来,[/font][font=Calibri]Detergent[/font][font=宋体]作为一种两亲性分子,疏水尾部包裹目的蛋白的疏水区域,亲水头部位于与溶液接触的界面。微团的形成是膜蛋白增溶的基础,当去垢剂浓度高于[/font][font=Calibri]CMC[/font][font=宋体]([/font][font=Calibri]Critical micelle concentration[/font][font=宋体],临界胶束浓度)时会形成微团,增溶后,去垢剂将蛋白周围的磷脂置换,从而实现收集目标膜蛋白的目的,后续再进行蛋白纯化,最终蛋白呈现在含有[/font][font=Calibri]Detergent[/font][font=宋体]的溶液中。义翘神州成功搭建了去垢剂技术平台,利用该平台可有效提高跨膜蛋白的产量和纯度。[/font][/font][font=宋体] [/font][b][font=宋体][font=Calibri]Nanodisc[/font][font=宋体]技术平台[/font][/font][/b][font=宋体][font=Calibri]Nanodisc[/font][font=宋体]结构稳定,与天然的生物膜非常相似,使得[/font][font=Calibri]Nanodisc[/font][font=宋体]能够很好地应用于膜蛋白的研究。目前[/font][font=Calibri]Nanodisc[/font][font=宋体]平台有[/font][font=Calibri]2[/font][font=宋体]种方式,一种是基于苯乙烯马来酸酐共聚物([/font][font=Calibri]SMA[/font][font=宋体])组装的[/font][font=Calibri]SMA-Nanodisc[/font][font=宋体]平台,如下图(左)所示,它可以直接从细胞膜上提取膜蛋白,使其变为可溶性蛋白,组装完成的蛋白样品很稳定,更能维持蛋白的天然构象。另一种是基于膜骨架蛋白([/font][font=Calibri]MSP[/font][font=宋体])的[/font][font=Calibri]MSP-Nanodisc[/font][font=宋体]平台(下图右),它需要先将膜蛋白利用去垢剂制备出来,然后再加入磷脂分子和[/font][font=Calibri]MSP[/font][font=宋体]进行组装。通过调整磷脂、[/font][font=Calibri]MSP[/font][font=宋体]和待组装膜蛋白三者的比例,可以使得待组装膜蛋白在[/font][font=Calibri]Nanodisc[/font][font=宋体]中呈不同聚集状态。义翘神州已成功搭建了[/font][font=Calibri]Nanodisc[/font][font=宋体]技术平台,利用跨膜蛋白与磷脂结合能够维持其良好活性的特性,制备出稳定的产品,满足动物免疫、抗体筛选、[/font][font=Calibri]cell-based assays[/font][font=宋体]等场景。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/transmembrane-proteins[/font][/font]
[size=15px][font=宋体]结直肠癌([/font][font=&]Colorectal Cancer[/font][font=宋体],[/font][font=&]CRC[/font][font=宋体])是全球常见的恶性肿瘤,术后复发转移已成为结直肠癌患者死亡的主要原因,特别是结直肠癌肝转移([/font][font=Georgia, &][color=#333333]CRLM[/color][/font][font=宋体])。因此,迫切需要深入研究[/font][font=Georgia, &][color=#333333]CRLM[/color][/font][font=宋体]的病理和分子机制。原发性肿瘤细胞与远端器官中的免疫细胞或基质细胞之间的信息传递是转移前微环境([/font][font=&]PMN[/font][font=宋体])形成的关键因素,了解这一机制对于制定有效的肿瘤转移治疗策略至关重要。[/font][font=&][/font][/size][size=15px][font=宋体]细胞外囊泡([/font][font=&]EV[/font][font=宋体])作为各种细胞分泌的功能实体,富含蛋白质、核酸、脂质和其他分子,促进肿瘤细胞和基质细胞之间的重要通讯。越来越多的报道表明,肿瘤衍生的[/font][font=&]EV[/font][font=宋体]([/font][font=&]TEV[/font][font=宋体])有助于[/font][font=&]PMN[/font][color=#333333]和[/color][font=Georgia, &][color=#333333]CRLM[/color][/font][font=宋体]的形成,为转移器官中循环肿瘤细胞的增殖提供必需肿瘤微环境([/font][font=&]TME[/font][font=宋体])。作者推测[/font][font=&]circRNA[/font][font=宋体]富集的[/font][font=&]TEVs[/font][font=宋体]介导[/font][font=&]PMN[/font][font=宋体]的形成,并且靶向[/font][font=&]circRNA[/font][font=宋体]富集的[/font][font=&]TEVs[/font][font=宋体]可能是针对[/font][font=&]PMN[/font][font=宋体]形成和[/font][font=&]CRLM [/font][font=宋体]的有效治疗策略。[/font][/size][size=15px][font=宋体]肿瘤细胞分泌的富含[/font][font=&]circ-0034880[/font][font=宋体]的细胞外囊泡通过增强[/font][font=&] SPP1[sup]high[/sup]CD206[sup] +[/sup][/font][font=宋体]促肿瘤巨噬细胞的活化来促进结直肠癌肝转移。重要的是,研究确定人参皂苷[/font][font=&]Rb1[/font][font=宋体]是一种潜在的治疗剂,通过直接靶向[/font][font=&]QKI [/font][font=宋体]蛋白,从而减少[/font][font=&]circ-0034880[/font][font=宋体]的生物合成并抑制[/font][font=&]SPP1[sup]high[/sup]CD206[sup] +[/sup][/font][font=宋体]促肿瘤巨噬细胞的活化,最终抑制结直肠癌肝转移。研究从调控[/font][font=&]TME[/font][font=宋体]的角度,特别是抑制[/font][font=&]circ-0034880[/font][font=宋体]的生物合成和细胞外囊泡的生成,首次揭示了人参皂苷[/font][font=&]Rb1[/font][font=宋体]在肿瘤防治领域的特殊作用,为今后的临床药物转化奠定了坚实的基础。 [size=15px][b]1、富含Circ-0034880的血浆EV与CRLM相关[/b][/size][font=宋体]基于细胞外囊泡在肿瘤转移中的重要作用,作[/font][font=宋体]者首先人类血液外泌体数据库、GSE159669数据集,以及临床血浆细胞外囊泡样本发现[/font][size=15px]一个环状RNA circ-0034880在结直肠癌肝转移患者中具有更高的表达水平,表明该环状RNA与结直肠癌的肝转移密切相关[/size] [size=15px][b]2、富含Circ-0034880的TEV在体内促进CRLM[/b][/size][size=15px]为了进一步研究circ-0034880富集的TEVs对CRC肝转移的体内影响,作者通过体内示踪实验发现了肿瘤细胞MC38来源的EVs在肝中高度积累。体内注射肿瘤细胞MC38来源的EVs可以促进肝转移,而缺失circ-0034880的EVs却丧失其促肝转移的作用,结果表明EVs依赖于其携带的circ-0034880发挥作用[/size] [size=15px][b]3、Circ-0034880富集的TEV激活肝脏转移前微环境中的CD206+促肿瘤巨噬细胞[/b][/size][size=15px]为了全面评估circ-0034880富集的TEV对肝脏转移前微环境的影响,研究人员对小鼠持续进行TEV注射和肿瘤细胞注射,建立了一个肝转移小鼠模型,多重免疫荧光分析显示TEV处理促进CD206 +促肿瘤巨噬细胞在转移前肝脏中的显著浸润,且circ-0034880表达水平与CD206 +促肿瘤TAM浸润之间存在正相关性 [/size][size=15px][/size][size=15px][b]4、Circ-0034880富集的TEV通过激活CD206 +促肿瘤巨噬细胞促进CRC细胞迁移[/b][/size][size=15px]由于有多项报道显示活化的巨噬细胞对CRC细胞迁移有促进作用,作者进一步研究利用示踪实验发现TEV可以携带circ-0034880被巨噬细胞所吸收。此外,功能实验表明富含circ-0034880 的TEV可以促进CD206+促肿瘤巨噬细胞的活化,并且携带circ-0034880的TEV的巨噬细胞上清会显著促进肿瘤细胞的迁移 [/size][size=15px][b]5、Circ-0034880富集的TEV促进SPP1[sup]high[/sup]CD206 +促肿瘤巨噬细胞的激活[/b][/size][size=15px]为了探究携带circ-0034880的TEV对CD206+促肿瘤巨噬细胞的激活机制,研究人员对TEV处理的巨噬细胞进行了转录组数据分析,进一步通过体外基因和蛋白检测和体内IF实验验证了巨噬细胞中SPP1是其调控的靶点,这些发现表明circ-0034880富集的TEV促进了SPP1[sup]high[/sup] CD206 +促肿瘤巨噬细胞的激活。鉴于已知circRNA可作为miRNA海绵发挥作用,作者分析了circ-0034880靶向的miRNA和SPP1结合的miRNA,发现了有2个miRNA重叠:miR-200a-3p和miR-141-3p。结合实验证明这两个miRNA可结合circ-0034880和SPP1,表明 circ-0034880和SPP1竞争结合miRNA,使SPP1不被miRNA所抑制。SPP1是巨噬细胞活化的关键蛋白,因此,研究结果表明TEV释放的circ-0034880通过保护SPP1免受miR-200a-3p和miR-141-3p介导的降解来提高巨噬细胞中SPP1的表达,促进SPP1[sup]high[/sup]巨噬细胞亚群增加 [/size][size=15px][b]6、人参皂苷Rb1给药可通过阻止富含circ-0034880的TEVs介导SPP1[sup]high[/sup]CD206+促肿瘤巨噬细胞的激活来阻止CRC细胞迁移[/b][/size][size=15px]鉴于circ-0034880的重要作用,下调其表达可以作为抑制癌症肝转移的策略,于是作者对103种天然药物进行了筛选,发现4种天然产物(人参皂苷Rb1、异鼠李素、山奈酚和槲皮素)对该circ-0034880的抑制作用最强,其中人参皂苷Rb1具有更强抑制作用。同样人参皂苷Rb1预处理的肿瘤细胞来源的TEV,其作用于巨噬细胞后下游的SPP1的蛋白表达造成下调作用,后续对结直肠癌细胞迁移能力下降,效果类似于沉默circ-0034880。总之,研究结果表明人参皂苷Rb1给药可通过抑制富含circ-0034880的TEV介导的SPP1[sup]high[/sup] CD206+促肿瘤巨噬细胞的激活来阻止CRC细胞迁移 [/size][size=15px][b]7、人参皂苷Rb1直接与QKI蛋白结合,抑制circ-0034880的生物合成[/b][/size][size=15px]为了确定影响circ-0034880表达的Rb1的直接靶点,作者进行了DARTS实验筛选出151种差异表达蛋白,其中一个蛋白QKI被报道与调控前mRNA剪接和促进circRNA生物合成。研究者接下来采用CETSA分析来验证了QKI和Rb1的结合,证实了Rb1能够显著增加QKI的热稳定性。进一步采用SPR分析验证了Rb1与QKI之间的很强的结合亲和力,通过分子对接预测了结合模式。此外,敲低QKI能够显著抑制该circRNA的表达。研究结果表明Rb1直接与QKI蛋白结合,抑制circ-0034880的生物合成[/size] [size=15px][/size][size=15px][b]8、[/b][/size][size=15px][b]人参皂苷[/b][/size][size=15px][b]Rb1给药通过阻止circ-0034880富集的TEV介导SPP1[sup]high[/sup]CD206+促肿瘤TAM的激活来抑制CRLM[/b][/size][size=15px]最后,作者验证了Rb1 给药的[i]体内[/i]效果,发现与单独使用TEV相比,使用Rb1预处理的TEV给药组的肝转移显著减少,与直接沉默circ-0034880的效果非常相似。然而,在沉默circ-0034880的情况下,使用Rb1预处理的TEV给药对肝转移的影响很小。接下来,作者探讨了Rb1对肝转移中CD206+促肿瘤TAM浸润的影响,发现在Rb1预处理的TEVs给药组中,SPP1表达显著下调,类似于直接沉默circ-0034880的效果。然而,在沉默circ-0034880的前提下,相应肝转移中的SPP1表达受到Rb1预处理的TEVs给药的影响最小。总之,体内功能实验证明Rb1预处理的肿瘤细胞来源的TEV失去了促进癌症肝转移的作用[/size][/font][/size]
鸡蛋,在如今的社会里,更多时候是作为一种营养丰富的食品出现在我们的餐桌上。现代化大型养殖场如生产产品般输出鸡蛋的方式颠覆了人们对鸡蛋的认识,或许已经很少有人能够联想到从蛋黄蛋白到一个小生命的奇迹升华。但在人类漫长的历史中,农业是文明的核心。就在不太遥远的过去,大多数人还可以在家中目睹鸡生蛋、蛋生鸡的奇迹。这种神秘的现象让古时的人们感到好奇、困惑,甚至产生莫名的崇拜。我们华夏文明由雏鸡的诞生联想到世界的起源,“天地混沌如鸡子,盘古生其中”,看来在我们祖先的眼中,鸡蛋的孵化犹如天地诞生般神秘。这种“卵生崇拜”在史籍中屡见不鲜,如《史记·殷本纪》记述商朝人先祖契的来历时提到有娀氏的女儿简狄“见玄鸟坠其卵,简狄取吞之,因孕生契”,同样,在《史记·秦本纪》中,文章伊始就记载了颛顼的孙女女修织布时“玄鸟陨卵,女修吞之,生子大业”,而这位大业就是秦人的先祖。不得不佩服古人的想象,这玄鸟蛋孵化出了两个重要朝代。在漫长的历史中,这种对蛋朦胧而浪漫的崇拜逐渐融入了我们的文化中,直到如今,染红壳的鸡蛋依旧是新婚、生子、满月时,人们表达祝福的重要载体。随着科技的进步,人们对蛋的理解逐渐清晰,现在很多人都知道蛋和卵细胞有千丝万缕的关系。可是鸡蛋到底是否就是一个细胞?答案可谓五花八门,有人说整个鸡蛋就是一个放大的卵细胞,蛋壳内的那层膜是细胞膜,蛋清是细胞质,蛋黄是细胞核;也有人说蛋黄是卵细胞,卵黄膜就是细胞膜,蛋黄就是细胞质,而蛋黄上面的小白点是细胞核;还有人认为鸡蛋本就是由很多细胞构成的。
功能性蛋白及一例分析自19世纪中叶荷兰化学家Gerardus Mul-der从动物组织和植物体中提取出蛋白质以来,人们发现了越来越多的蛋白质,据估计生物界中蛋白质的种类可达1010~1012之多;在这如此众多的蛋白质中,功能性蛋白发挥着极其重要的生理功能 。功能性蛋白也有人称其为活性蛋白。它们的特点是都有识别功能,能与其他分子特异性结合.完成各种复杂的生命活动:在结构上主要是一些球状蛋白质。1 功能性蛋白的种类按其作用方式不同可分为酶蛋白、运输蛋白、运动蛋白、免疫球蛋白、毒蛋白、激素蛋白(1)酶蛋白: 细胞的生长和繁殖、代谢物的合成和分解、能量的产生和利用,这些过程所需要的物质都是通过无数的生物化学反应来提供的.而这些反应又都是在一类特殊蛋白质—酶蛋白的催化下完成的。酶的催化效率极高,且具有高度的专一性,也正是这种高度的专一性使一种特定的酶只能作用于一种或少数几种结构相似的化合物,这就要求有各种不同的酶去作用于不同的化合物。在酶的作用下,生物细胞才得以合成各种复杂的化合物,也才能使各种大分子物质被分解、吸收和利用.且这些反应都要在适合于生物体本身的温度、压力和pH值等非常温和的条件下进行,能使生物细胞按照这种方式进行化学变化是蛋白质最重要的功能之一。常见的酶蛋白如淀粉酶使淀粉分解形成葡萄糖,蛋白酶、肽酶使蛋白质分解为氨基酸;溶菌酶使细菌细胞壁中的肤聚糖被破坏;凝血系统酶的有序作用使凝血过程得以有条不紊地进行.合成酶能合成多种体内所需要的大分子物质。应用举例:由于近年来鱼粉资源价格上涨,冷向军等人通过向鱼粉含量较低(10﹪)的饲料个添加蛋白酶AG使鱼的前肠蛋白酶有显著提高。同样有实验证明在玉米-豆粕型粮食中添加蛋白酶可以改善肉鸡的生长性能,提高蛋白质的消化率。(2)运输蛋白:有些蛋白质起载体的作用可以运输特定的物质到达必须的部位,使其完成特定的功能,这种蛋白质称为运输蛋白。如哺乳动物的血红蛋白能将氧从氧气充裕的肺内运送到各个组织中去:血清蛋白能与游离脂肪酶等多种物质结合,并将这些物质在脂肪组织与身体的其他部位间运送(最典型的β1-脂蛋白可随血流运输脂肪),铁传递蛋白能传递血液中的铁。无脊椎动物体内的血蓝蛋白,大豆根瘤中的豆血红蛋白也起着输送氧气的作用。另外还有一些能携带物质通过细胞膜进出细胞的蛋白质,如细菌过膜运输中的载体蛋白等,它们都属于运输蛋白。(3)运动蛋白:参与运动功能的蛋白质种类较多如脊椎动物中骨骼肌的主要成分就是肌动蛋白和肌球蛋白,肌肉的收缩就是靠着这两种互相联系的平行丝状蛋白相对滑动来完成的;细菌的运动器官——鞭毛也是由鞭毛蛋白组成的;绿藻的运动也离不开蛋白质;有丝分裂的完成,精子的运动等都与运动蛋白有关,所以绝大多数生物的运动和收缩过程都是运动蛋白参与的结果。应用举例:邱永忠等人在研究烟草花叶病毒(TMV)在植物细胞间的运动时发现用体外定位突变引起L株上,被点突变的DNA体外转录成RNA后感染感病烟草,结果定位突变的L株表型30kD蛋白基因四种位点不同的移码突变和一种基因中间大部分缺失的突变体均使病毒不能感染植株。这证明TMV 30kD蛋白与病毒运动有关,而与病毒复制无关。同时因为胞间连丝一般只能让小于1kD的分子通过,其通透范围远小于病毒颗粒,也小于折叠的病毒核酸分子,Wolf等实验证明正时因为30kD蛋白才使得植株分子半径扩散了三倍多。(4)免疫球蛋白:指具有抗体活性的动物蛋白。主要存在于血浆中,也见于其他体液、组织和一些分泌液中。脊椎动物的免疫系统能抵抗外来的入侵物质,如病毒、细菌以及其他机体的细胞,当外来的这些入侵物质(抗原)进入机体后就会激发机体的免疫系统而产生特异性的免疫球蛋白(抗体),通常每一种抗体对于相应的某一特定抗原具有高度的专一性,抗原与抗体结合形成抗原-抗体复合物.使入侵物质——抗原失活而排出体外,从而消除外来物质对机体的干扰。由此看来蛋白质不仅参与了高等动物的免疫反应,而且起着重要的作用,由于抗原和抗体结合的高度专一性,必然有数量众多的抗体作用于不同的抗原物质,据估计抗体的类型可能有10O万种,即免疫球蛋白可能有100万种之多。(5)毒蛋白:动物、植物和微生物都可以产生某些特殊的物质来防御敌害,这些物质中绝大多数是蛋白质类物质,由于它们对高等动物具有毒性,故称为毒蛋白。蝎类能产生毒性很强的蝎毒蛋白.用来攻击敌害,保护自己;蛇类产生的神经毒素和心赃毒素其主要成分也是小分子量的蛋白质;毒蘑菇中的相当一部分蘑菇毒素也是蛋白质;细菌产生的毒素,毒性极强的肉毒梭菌毒素(人的致死量小于19m)和破伤风痉挛毒素、白喉杆菌毒素等外毒素均是蛋白质。应用举例:王峰等人研究核糖体失活蛋白(RIPS)是一类能够抑制细胞核糖体合成蛋白质,从而导致宿主死亡的毒蛋白,广泛存在于植物、细菌中。发现其在在细胞内的转运途径研究很多,目前较为清楚的是逆向转运途径,其中以蓖麻毒素、志贺菌毒素、霍乱毒素为代表,大体过程为:内吞一内吞小体一高尔基体一内质网一胞液。(6)激素蛋白:是由特殊细胞所产生的一类物质,它们通过与靶细胞或系统内其它器官的相互作用来发挥其代谢上的功能,其实许多激素本身就是蛋白质,这样的蛋白质称为激素蛋白,它们在生物合成上具有重要的功能。如胰高血糖素、胰岛素、胃泌素、生长激素、促甲状腺激素、促肾上腺皮质激素和促脂解激素等均是蛋白
[font=宋体][font=宋体]膜联蛋白([/font][font=Calibri]Annexin[/font][font=宋体])是一类分布广泛的钙依赖性磷脂结合蛋白,与磷脂酰丝氨酸([/font][font=Calibri]PS[/font][font=宋体])能特异性结合,参与一系列[/font][font=Calibri]Ca2+[/font][font=宋体]依赖型的膜相关的过程,包括细胞的胞吐和内吞作用、囊泡运输、调节血液凝固以及炎症反应等多种生物学事件,在许多人类疾病的发病机制或进展中起着非常重要的作用。膜联蛋白[/font][font=Calibri]V[/font][font=宋体]([/font][font=Calibri]Annexin V[/font][font=宋体])染色是检测细胞凋亡的常用方法。[/font][/font][b][font=宋体][font=宋体]膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色原理及应用[/font][/font][/b][font=宋体] [/font][b][font=宋体][font=宋体]一、膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色原理[/font][/font][/b][font=宋体] [/font][font=宋体][font=宋体]膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色,也称为[/font][font=Calibri]Annexin V[/font][font=宋体]染色,是一种用于检测细胞凋亡的方法。其核心原理基于细胞凋亡过程中的一种生物化学变化。在正常细胞中,磷脂酰丝氨酸([/font][font=Calibri]PS[/font][font=宋体])只分布在细胞膜脂质双层的内侧。然而,当细胞开始凋亡时,这一分布会发生改变,磷脂酰丝氨酸会从细胞膜内侧翻到外侧。膜联蛋白[/font][font=Calibri]V[/font][font=宋体]是一种能够与这种外翻的磷脂酰丝氨酸特异性结合的蛋白。通过结合荧光物质,这种结合可以被检测和观察,从而确定哪些细胞正在经历凋亡。[/font][/font][b][font=宋体] [/font][font=宋体][font=宋体]二、膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色的应用[/font][/font][/b][font=宋体] [/font][font=宋体][font=宋体]①流式细胞术:膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色常用于流式细胞术中,以检测和分类正常细胞和凋亡细胞。通过流式细胞仪,可以快速分析大量细胞,并准确地识别出凋亡细胞。[/font][/font][font=宋体][font=宋体]②光学显微镜成像:膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色也可用于光学显微镜成像技术,这使得研究者能够在显微镜下直接观察细胞的形态变化,从而对凋亡过程有更深入的理解。[/font][/font][font=宋体][font=宋体]③与其他染色方法的结合:膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色可以与其他染色方法如碘化丙啶[/font][font=Calibri](PI)[/font][font=宋体]染色结合使用。[/font][font=Calibri]PI[/font][font=宋体]是一种能够进入凋亡晚期细胞核的染料,因此可以用于区分凋亡早期和晚期细胞。这种联合使用的方法能提供更全面的细胞凋亡信息。[/font][/font][font=宋体][font=宋体]④临床应用:膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色在许多临床领域中都有应用,例如肿瘤学、血液学和药理学等。它可以帮助研究者深入理解疾病的发展过程,评估新药物对细胞凋亡的影响,以及监测疾病的进展和治疗的效果。[/font][/font][font=宋体][font=宋体]总的来说,膜联蛋白[/font][font=Calibri]V[/font][font=宋体]染色是一种强大的工具,可以帮助科学家们更好地理解细胞凋亡的过程,从而为疾病的治疗和药物研发提供有价值的信息。[/font][/font][font=宋体] [/font][font=宋体]更多关于膜联蛋白详情可以关注[url=https://cn.sinobiological.com/][b]义翘神州[/b][/url]![/font][font=宋体] [/font][b][font=宋体][font=宋体]义翘神州:蛋白与抗体的专业引领者,欢迎通过百度搜索[/font][font=宋体]“义翘神州”与我们取得联系。[/font][/font][/b]
[font=宋体][b]什么是重组蛋白?[/b][/font][font=宋体] [/font][font=宋体][font=宋体][url=https://cn.sinobiological.com/resource/protein-review/protein-production][b]重组蛋白[/b][/url]的产生是应用了重组[/font][font=Calibri]DNA[/font][font=宋体]或重组[/font][font=Calibri]RNA[/font][font=宋体]的技术从而获得的蛋白质。体外重组蛋白的生产主要包括四大系统:原核蛋白表达,哺乳动物细胞蛋白表达,酵母蛋白表达及昆虫细胞蛋白表达。生产的蛋白在活性和应用方法方面均有所不同。根据自身的下游运用选择合适的蛋白表达系统,提高表达成功率。[/font][/font][font=宋体] [/font][font=宋体][b]融合蛋白和重组蛋白的区别[/b][/font][font=宋体][font=Calibri]1[/font][font=宋体]、重组蛋白[/font][/font][font=宋体]重组蛋白是指应用基因重组技术,获得连接有可以翻译成目的蛋白的基因片段的重组载体,之后将其转入可以表达目的蛋白的宿主细胞从而表达特定的重组蛋白分子。融合蛋白表达只是重组蛋白表达的一种策略,融合表达是一种方法。因为融合表达具有表达效率高、产物稳定而且水溶性好、易于鉴定和纯化等优点,现已被广泛采用。[/font][font=宋体][font=Calibri]2. [/font][font=宋体]融合蛋白[/font][/font][font=宋体][font=宋体]融合蛋白是指通过重组[/font][font=Calibri]DNA[/font][font=宋体]技术将你要表达的目的蛋白基因同表达载体上融合蛋白基因相连,这样表达出的蛋白质就会是同时具有目的基因蛋白和融合基因蛋白两个部分的重组蛋白。[/font][/font][font=宋体][font=宋体]融合蛋白与重组蛋白不是一个层次上对立的概念,融合蛋白表达只是重组蛋白表达的一种策略,融合表达是一种方法。因为融合表达具有表达效率高、产物稳定而且水溶性好、易于鉴定和纯化等优点,现已被广泛采用。融合蛋白又称标签([/font][font=Calibri]Tag[/font][font=宋体]),常用的[/font][font=Calibri]His[/font][font=宋体]、[/font][font=Calibri]GST[/font][font=宋体]等。[/font][/font][font=宋体] [/font][font=宋体]总结:在生物制药领域,重组蛋白具有较高的活性和纯度,更易吸收,安全性也更高的特点。重组蛋白的利用率也更高。[/font][font=宋体] [/font][font=宋体]为了生产重组蛋白,基因被分离并克隆到表达载体中。重组蛋白的生产需要蛋白表达系统、蛋白纯化系统和蛋白识别系统。[/font][font=宋体] [/font][font=宋体][b]获取重组蛋白的基本步骤:[/b][/font][font=宋体] [/font][font=宋体][font=Calibri]1.[/font][font=宋体]目标基因的扩增。[/font][/font][font=宋体][font=Calibri]2.[/font][font=宋体]插入克隆载体。[/font][/font][font=宋体][font=Calibri]3.[/font][font=宋体]亚克隆到表达载体中。[/font][/font][font=宋体][font=Calibri]4.[/font][font=宋体]转化到蛋白表达宿主中[/font][font=Calibri]([/font][font=宋体]细菌[/font][font=Calibri]([/font][font=宋体]大肠杆菌[/font][font=Calibri])[/font][font=宋体]、酵母细胞、哺乳动物细胞或杆状病毒[/font][font=Calibri]-[/font][font=宋体]昆虫细胞系统[/font][font=Calibri])[/font][font=宋体]。[/font][/font][font=宋体][font=Calibri]5.[/font][font=宋体]重组蛋白鉴定试验[/font][font=Calibri](Western blot[/font][font=宋体]或荧光[/font][font=Calibri])[/font][/font][font=宋体][font=Calibri]6.[/font][font=宋体]大规模生产。[/font][font=Calibri]([/font][font=宋体]大规模发酵[/font][font=Calibri])[/font][/font][font=宋体][font=Calibri]7.[/font][font=宋体]分离和纯化。[/font][/font][font=宋体] [/font][font=宋体]需要考虑多种因素:[/font][font=宋体][font=Calibri]1.[/font][font=宋体]选择哪个宿主系统?[/font][/font][font=宋体][font=Calibri]2.[/font][font=宋体]如何分离和纯化重组蛋白?[/font][/font][font=宋体] [/font][font=宋体]选择适当的表达宿主或使用正确的纯化方法并不容易,应考虑目标重组蛋白的性质。下面列出了一些重要因素:[/font][font=宋体] [/font][font=宋体]? 膜结合[/font][font=宋体]? 溶解度[/font][font=宋体]? 单或多结构域[/font][font=宋体][font=宋体]? 大小[/font][font=Calibri]([/font][font=宋体]分子量[/font][font=Calibri])[/font][/font][font=宋体]? 表达位置[/font][font=宋体] [/font][font=宋体][font=宋体]对于大多数没有足够经验来表达和分离重组蛋白的人来说,重组蛋白的生产是非常耗时的。许多生物公司为各种不同规模的重组蛋白表达提供良好的服务,例如义翘神州[/font][font=Calibri]([/font][font=宋体]参考重组蛋白生产的详细服务清单)[/font][/font][font=宋体] [/font][font=宋体]义翘神州提供重组蛋白和[url=https://cn.sinobiological.com/resource/protein-review/fusion-protein][b]融合蛋白[/b][/url]等相关信息,详情可以关注[/font][font=宋体][font=宋体]融合蛋白:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/fusion-protein[/font][/font][font=宋体] [/font][font=宋体][font=宋体]重组蛋白生产:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/protein-production[/font][/font]
1994年,Marc Wilkins在Siena双向凝胶电泳(two-dimensional electrophoresis,2-DE)会议上最早提出了蛋白质组(proteome)概念,并于1995年7月的Electrophoresis杂志上发表。随着高通量、高灵敏度、高分辨率生物质谱技术的出现,蛋白质组学技术取得飞速发展,人们不再满足于对一个细胞或组织的蛋白质进行定性研究,而是着眼于蛋白质量的研究,于是蛋白质组学概念就被提出,并得到了广泛的应用。蛋白质组学(Proteomics)是蛋白质(protein)与 基因组学(genomics)两个词的组合体,表示“一种基因组所表达的全套蛋白质”,即包括一种细胞乃至一种生物所表达的全部蛋白质。蛋白质组学研究,就是要把一个基因组表达的绝大多数蛋白质或一个复杂的混合体系中绝大多数蛋白质进行精确的定量和鉴定。蛋白质组本质上指的是在大规模水平上研究蛋白质的特征,包括蛋白质的表达水平,翻译后的修饰,蛋白与蛋白相互作用等,由此获得蛋白质水平上的关于疾病发生,细胞代谢等过程的整体而全面的认识。蛋白质组学是一门以全面的蛋白质性质研究为基础,在蛋白质水平对疾病机理、细胞模式、功能联系等方面进行探索的科学。目前最新的iTRAQ蛋白定量分析技术在此基础上被提出,并被得到广泛应用。仅仅知道蛋白质的身份并不足以对蛋白质给出最终定论,因为蛋白质的浓度对于实现其在细胞中的功能来说极其重要,一种特殊蛋白质在浓度上的变化,就能预示细胞的突变过程。因此,科学家能够对蛋白质的相对和绝对浓度进行测量,是很重要的事情。过去,科学家通常先进行二维(2D)凝较电泳,切断条带,再用质谱方法测量条带中的蛋白质。可是,这种方法不是很理想:既不是非常敏感,也不是非常精确。新泽西医学及牙科大学的蛋白组学研究中心主任Hong Li说:“当我们开始蛋白组学研究时,就采用2D凝胶技术,但得出的信息量却让大伙很失望,因为许多蛋白质已经改变了自身的代谢过程,如热休克蛋白或者是管家蛋白。”蛋白组学中的方法一直在不断提高。基于高度敏感性和精确性的串联质谱方法,不需要凝胶,就可以获得相对和绝对定量的蛋白质结果。iTRAQ和iCAT是这些新进展中的两大主力,但是,新技术也有不尽如人意的地方,需要不断改进。iTRAQ技术是由美国应用生物系统公司(Applied Biosystems Incorporation,ABI)2004年开发的同位素标记相对和绝对定量(isobaric tags for relative and absolute quantitation,iTRAQ)技术,是一种新的、功能强大的可同时对四种样品进行绝对和相对定量研究的方法,这种方法是建立在iTRAQ试剂的基础上。上海舜百生物公司目前所采用的就是这种iTRAQ技术。该技术的主要特点在于:1. 分离能力强、分析范围广;2. 定性分析结果可靠,可以同时给出每一个组分的分子量和丰富的结构信息;3. MS具备高灵敏度、检测限低;4. 分析时间快,分离效果好;5. 自动化程度高;6. iTRAQ技术不仅可发现胞浆蛋白,还有膜蛋白、核蛋白、胞外蛋白。iTRAQ技术可检测出低丰度蛋白、强碱性蛋白、小于10KD或大于200KD的蛋白。上海舜百生物使用的液相色谱仪的型号是日本岛津公司2D-nano-HPLC,质谱仪型号是美国ABI公司的MALDI-TOF-TOF 4700,标记试剂盒是美国ABI公司的ITRAQ标记试剂盒。舜田生物所采用的iTRAQ试剂是一种小分子同重元素化学物质,包括三部分:一端是报告部分(reporter group ),另一端是肽反应部分(peptide reactive group),中间部分是平衡部分(balance group)。 其中,reporter group:质量为114Da、115 Da、116 Da、117 Da,因此iTRAQ试剂共四种。 peptide reactive group:将reporter group与肽N端及赖氨酸侧链连接,几乎可以标记样本中所有蛋白质。 balance group:质量为31 Da、30 Da、29 Da、28 Da,使得四种iTRAQ试剂分子量均为145 Da,保证iTRAQ标记的同一肽段m/z相同。舜百生物公司iTRAQ的操作程序如下:将蛋白质裂解为肽段,然后用iTRAQ试剂进行差异标记。再将标记的样本相混合,这样就可以对其进行比较。与样本结合后,通常用MudPIT多维蛋白质鉴定技术进行下一步的操作,用2D液相色谱串联质谱进行分析。在质谱分析鉴定特殊肽离子片断结构的基础上,采用美国应用生物系统公司的软件包MASCOT和Protein Pilot对每一个肽段进行鉴定。其具体操作如下图所示: http://img.dxycdn.com/trademd/upload/userfiles/image/2013/11/A1383890263png_small.jpgiTRAQ技术对检测标本也有一定要求。舜百生物要求检测蛋白量最低不少于50 ug,浓度最低不少于5 ug/uL,否则同位素无法标记。而对蛋白提取试剂也要求使用普通的组织、细胞裂解液即可,切忌不要使用二维电泳试剂提取。iTRAQ技术区别与以往二维电泳技术具有更明显的优势,两者比较如下:1. 二维电泳所检测的发生表达变化的蛋白都位于细胞浆内,而iTRAQ可检测到蛋白有胞浆蛋白外,还有线粒体蛋白、膜蛋白和核蛋白。2. 二维电泳观察到的蛋白变化在2倍以上,而用iTRAQ计算出的蛋白变化在1.3-1.6倍之间。3. iTRAQ技术在鉴定大分子和小分子蛋白方面也有优势。4. 二维电泳是通过切断条带,再用质谱方法测量条带中的蛋白质,但该方法既不是非常敏感,也不是非常精确,获取的信息量很少。 而itraq技术是基于高度敏感性和精确性的串联质谱方法,不需要凝胶,就可以获得相对和绝对定量的蛋白质结果。5. iTRAQ可以对任何类型的蛋白质进行鉴定,包括高分子量蛋白质、酸性蛋白质和碱性蛋白质,而二维电泳对这些蛋白质都束手无策。由此,舜百生物得出结论:iTRAQ技术比二维电泳技术能发现更多数量和种类的蛋白,但在比较胞浆蛋白及蛋白量的变化方面,二维电泳技术有一定长处,对iTRAQ的实验结果有互补作用。
[font=宋体][font=宋体]泛素化是一种细胞内的蛋白质标记系统,蛋白质泛素化是指将小的蛋白质泛素共价地连接到其他蛋白质分子上的过程。泛素([/font][font=Calibri]ubiquitin[/font][font=宋体])是一种高度保守的蛋白质,其结构由[/font][font=Calibri]76[/font][font=宋体]个氨基酸残基组成。泛素连接到目标蛋白质上的过程,经历了泛素激活、泛素转移和靶蛋白接受三个主要步骤。[/font][/font][font=宋体] [/font][font=宋体]蛋白质泛素化具有多种特点,例如它是高度选择性的,不同蛋白质泛素化的位置和数量可以影响其功能;它是可逆的,通过去泛素化反应可以调控蛋白质的泛素化状态;它还是动态调控的,受到多种因素的调控,如细胞信号通路和环境刺激。[/font][b][font=宋体]泛素化蛋白大小:[/font][/b][font=宋体] [/font][font=宋体][font=宋体]蛋白泛素化是指将小蛋白颗粒泛素([/font][font=Calibri]Ubiquitin[/font][font=宋体])与其他蛋白质共价结合的修饰过程。 泛素化修饰通常会导致泛素共价连接在蛋白质的赖氨酸残基上形成多重泛素链。 这种蛋白质泛素化增加了蛋白质的分子量,因为每个泛素分子的质量大约为[/font][b][font=Calibri]8.5[/font][font=宋体]千达尔顿([/font][font=Calibri]kDa[/font][/b][font=宋体][b])[/b]。[/font][/font][font=宋体] [/font][font=宋体] [/font][b][font=宋体]泛素化蛋白质组学在许多领域有重要的应用,主要包括:[/font][/b][font=宋体] [/font][font=宋体]①疾病机制研究:泛素化是一种广泛存在于细胞中的蛋白质修饰方式,参与了细胞的生长、分化、修复和调控等多个生命活动。泛素化蛋白质组学的研究可以帮助我们了解泛素化修饰的生物学功能和调控机制,为疾病发生机制和治疗策略的研究提供重要线索。例如,在癌症、代谢综合征、神经退行性疾病等疾病中,则会出现异常泛素化。[/font][font=宋体]②药物研发:通过分析药物对泛素化蛋白质的影响,可以评估药物的效力和选择性,为药物研发提供指导。[/font][font=宋体]③临床诊断:泛素化蛋白质组学鉴定与定量分析技术可以揭示细胞调控的机制,通过分析泛素化蛋白质的组学数据,可以确定泛素化修饰在细胞信号转导、蛋白质降解和细胞周期调控等过程中的重要作用。此外,通过比较病态和正常样品中泛素化蛋白质的差异,可以鉴定与疾病发生发展相关的泛素化修饰靶点,并进一步理解疾病的分子机制。因此,这些技术也可用于临床诊断。[/font][font=宋体]④蛋白质降解调控:在癌症、神经退行性疾病和免疫相关疾病等病症中,蛋白质降解调控出现异常。而泛素化蛋白组在调控蛋白质降解中发挥重要作用。通过与泛素连接,目标蛋白质被送入蛋白酶体或蛋白酶体样体中进行降解。这个过程是细胞清除异常、老化或受损蛋白质的重要途径。[/font][font=宋体]⑤高通量技术应用:高通量泛素化蛋白质组学鉴定与定量分析技术的发展包括质谱鉴定和抗体鉴定两种方法。质谱鉴定技术利用质谱仪的高灵敏度和分辨率,能够鉴定泛素化修饰的蛋白质及其泛素化位点。抗体鉴定技术则通过特异性抗体的使用,可以富集和鉴定泛素化修饰的蛋白质。这些技术为全面了解泛素化在细胞中的作用机制和调控网络提供了可能。[/font][font=宋体]总的来说,泛素化蛋白质组学在多个领域都有重要的应用价值,推动了我们对生命过程的深入理解以及疾病治疗的创新发展。[/font][font=宋体] [/font][font=宋体]更多详情关于[url=https://cn.sinobiological.com/resource/protein-review][b]蛋白资源[/b][/url]详情可以参看:[/font][url=https://cn.sinobiological.com/resource/protein-review][u][font=宋体][color=#0000ff][font=Calibri]https://cn.sinobiological.com/resource/protein-review[/font][/color][/font][/u][/url][b][font=宋体] [/font][font=宋体][font=宋体]义翘神州:蛋白与抗体的专业引领者,欢迎通过百度搜索[/font][font=宋体]“义翘神州”与我们取得联系。[/font][/font][/b]
[font=宋体][font=宋体]原核表达(这里是指[/font] [font=宋体]大肠杆菌表达系统[/font] [font=宋体])是目前常用的重组蛋白表达方式之一,操作相对简便,要求较低。相对哺乳动物表达系统更加经济节约。但是也常常遇到一些问题,比如原核表达蛋白表达量低?蛋白不表达?蛋白表达不在上清易形成包涵体等情况。本文主要根据实验经验总结[/font] [font=宋体]原核蛋白表达[/font] [font=宋体]的两个问题蛋白为什么不表达和蛋白表达为什么不在上清,并提出针对性的解决方案,帮助我们提高实验的成功率。原核表达[/font][font=Calibri]FAQ[/font][font=宋体]主要内容如下:[/font][/font][font=宋体] [/font][font=宋体]一、蛋白表达为什么不在上清?[/font][font=宋体] [/font][font=宋体][font=宋体]蛋白在大肠杆菌中的表达部位:细胞质中、细胞周质中、细胞外。[/font] [/font][font=宋体]蛋白在细胞周质中表达:细胞周质是指革兰氏阴性菌中、位于内膜和外膜之间的结构部分。周质中表达的蛋白质,分离纯化简单,而且周质中的氧化环境有利于蛋白质的正确折叠,但是蛋白产量很低。[/font][font=宋体]蛋白在胞外表达:胞外分泌是使大肠杆菌中的外源蛋白分泌到培养基中。[/font][font=宋体]途径:[/font][font=宋体]①用大肠杆菌细胞固有的途径,使真正属于分泌的蛋白直接分泌到胞外培养基中。大肠杆菌在正常情况下只有极少的蛋白可以分泌到胞外,且不是特别有效。[/font][font=宋体]②诱导大肠杆菌细胞的外膜发生有限的渗漏,从而使细胞内的蛋白向胞外培养基中分泌。[/font][font=宋体] [/font][font=宋体]二、蛋白的分泌表达优点[/font][font=宋体][font=Calibri]1[/font][font=宋体]、一些可被细胞内蛋白酶所降解的蛋白质分泌到周质或培养基中可增加其稳定性;[/font][/font][font=宋体][font=Calibri]2[/font][font=宋体]、有些在细胞内表达时无活性的蛋白分泌表达时能够按适当的方式进行准确折叠,增加到蛋白的活性;[/font][/font][font=宋体][font=Calibri]3[/font][font=宋体]、 由于蛋白质信号肽和编码序列间被切割,所以分泌蛋白产物不含氨基酸起始密码子[/font][font=Calibri]ATG[/font][font=宋体]所编码的甲硫氨酸,而甲硫氨酸会影响许多蛋白的活性。[/font][/font][font=宋体] [/font][font=宋体]三、如何提高蛋白的可溶性?[/font][font=宋体]重组蛋白在大肠杆菌中大量表达时,很容易在胞内形成不可溶的包涵体,为后期的纯化、复性带来麻烦,而且复性所得到的蛋白活性很低甚至没有活性。因此,提高重组蛋白在大肠杆菌中的可溶性表达是十分必要的。[/font][font=宋体] [/font][font=宋体]四、蛋白为什么不表达?[/font][font=宋体]真核基因在原核细胞中的的特点[/font][font=宋体]①使用原核启动子[/font][font=宋体][font=宋体]原核细胞[/font][font=Calibri]RNA[/font][font=宋体]聚合酶能识别真核基因启动子。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]②真核基因要置于原核载体[/font][font=Calibri]SD[/font][font=宋体]序列后[/font][/font][font=宋体][font=宋体]从真核[/font][font=Calibri]DNA[/font][font=宋体]转录的[/font][font=Calibri]mRNA[/font][font=宋体]缺乏结合原核核糖体的[/font][font=Calibri]SD[/font][font=宋体]序列,不能启动翻译过程。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]③使用[/font][font=Calibri]cDNA[/font][/font][font=宋体][font=宋体]真核基因中含有内含子,而原核细胞缺乏转录后加工体系,[/font][font=Calibri]mRNA[/font][font=宋体]中内含子不能切除,不能形成成熟的[/font][font=Calibri]mRNA[/font][font=宋体],也就不能表达真核蛋白。[/font][/font][font=宋体] [/font][font=宋体]④表达无需糖基化等修饰的蛋白[/font][font=宋体]原核细胞缺乏真核细胞所特有的蛋白翻译后的修饰系统,不利于表达需要糖基化、磷酸化修饰才有活性的真核蛋白。[/font][font=宋体] [/font][font=宋体]⑤真核基因必须切除信号肽序列[/font][font=宋体]蛋白具有信号肽序列的真核基因在真核细胞内先表达成蛋白前体,在其穿越细胞膜向外分泌时,会被细胞膜上信号肽识别序列切除而分泌到细胞外。但是在原核细胞内表达时,不但影响蛋白的表达,同时由于大肠杆菌缺乏加工处理体系,不能正确切除信号肽,而表达的分泌蛋白的前体没有实物活性,常聚集在胞内。[/font][font=宋体] [/font][font=宋体]五、影响原核表达的因素[/font][font=宋体][font=Calibri]1[/font][font=宋体]、翻译起始位点[/font][/font][font=宋体]现在大部分的表达载体都提供起始位点,所以它已经把起始密码子与核糖体结合位点的距离进行了优化,一般情况下不需要自己再添加,不过还是要留意载体图谱上是否注明有起始密码子和终止密码子。[/font][font=宋体] [/font][font=宋体][font=Calibri]2[/font][font=宋体]、[/font][font=Calibri]GC[/font][font=宋体]含量[/font][/font][font=宋体][font=宋体]表达序列中的[/font][font=Calibri]GC[/font][font=宋体]含量超过[/font][font=Calibri]70%[/font][font=宋体]的时候可能会降低蛋白在大肠杆菌中的表达水平。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3[/font][font=宋体]、[/font][font=Calibri]mRNA[/font][font=宋体]二级结构[/font][/font][font=宋体][font=宋体]在起始密码子附近的[/font][font=Calibri]mRNA[/font][font=宋体]二级结构可能会抑制翻译的起始或者造成翻译暂停从而产生不完全的蛋白。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4[/font][font=宋体]、密码子的偏爱性[/font][/font][font=宋体]如果外源基因密码子的使用频率和宿主菌高效表达的基因密码子的使用频率差异较大,翻译时核糖体在稀有密码子处就会产生停顿,这不仅降低蛋白质的合成效率,导致新生肽链的错误折叠而影响延伸,甚至会停止翻译。[/font][font=宋体] [/font][font=宋体][font=Calibri]5[/font][font=宋体]、质粒载体的选择[/font][/font][font=宋体]构建质粒载体时,要考虑质粒上的元件包括启动子,多克隆位点,终止密码子,融合标签,复制子,筛选标记基因等因素。[/font][font=宋体] [/font][font=宋体][font=Calibri]6[/font][font=宋体]、基因或者蛋白的大小[/font][/font][font=宋体][font=宋体]一般来说小于[/font][font=Calibri]10kd[/font][font=宋体]和大于[/font][font=Calibri]100kd[/font][font=宋体]的蛋白都是难以表达的。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]7[/font][font=宋体]、外源基因对宿主有毒性[/font][/font][font=宋体]外源基因在宿主内会抑制宿主菌的生长甚至导致宿主死亡,表观的现象:宿主菌在诱导后菌体量上升缓慢或不在生长。[/font][font=宋体] [/font][font=宋体][font=Calibri]8[/font][font=宋体]、基因突变(移筐突变)[/font][/font][font=宋体]碱基的突变、插入或缺失对阅读框的影响,可能会导致表达的蛋白质结构改变,或蛋白质合成过早终止。[/font][font=宋体] [/font][font=宋体][font=Calibri]9[/font][font=宋体]、培养诱导条件[/font][/font][font=宋体][font=宋体]培养基成分([/font][font=Calibri]C/N[/font][font=宋体])及培养条件(温度、诱导时机、诱导剂,浓度、诱导时间)也是影响蛋白表达的。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]更多关于[url=https://cn.sinobiological.com/resource/protein-review/protein-expression][b]蛋白表达[/b][/url]详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/resource/protein-review/protein-expression[/font][/font][font=Calibri] [/font]







 我要推广仪器
我要推广仪器
 下载APP
下载APP