[color=#444444]最近在做残留分析实验,咨询一下大师们关于农药苯噻酰草胺,苄嘧磺隆,吡嘧磺隆在高效液相色谱分析时最佳的色谱柱及条件?在这里先表示真挚的感谢![/color]
有同时测吡氟禾草灵和精吡氟禾草灵的吗?我们当时测的有点混了,如果有用MS测的请提供他们分别的离子,谢谢!吡氟禾草灵和精吡氟禾草灵的英文名称分别是fluazifop和fluazifop-p-butyl.
2009年10月7日,美国发布通报,美环保署拟制定杀虫剂Meptyldinocap、氯吡嘧磺隆、多杀菌素的许可限量。 本最终法规规定葡萄内/表Meptyldinocap,2-(1-methylheptyl)-4,6-dinitrophenyl (2E)-2-butenoate及按Meptyldinocap表示的2,4-DNOP, 2,4-dinitro-6-(1-methylheptyl)phenol混合残留的许可限量为:0.20 ppm。 本最终法规规定大豆种内的氯吡嘧磺隆(Halosulfuron-methyl)及其代谢物及降解物残留许可限量为:0.05 ppm。许可限量合格性将通过检测产品内/表氯吡嘧磺隆(Halosulfuron-methyl, methyl3-chloro-5-[[[[(4,6-dimethoxy-2-pyrimidinyl)amino]carbonyl]amino]sulfonyl]-1-methyl-1H-pyrazole-4-carboxylate来确定。 本最终法规规定以下作物内/表两种相关活性成分:Spinosyn A (Factor A CAS#131929-60-7)或2-[(6-deoxy-2,3,4-tri-O-methyl-[alpha]-L-manno-pyranosyl)oxy]-13-[[5-(dimethylamino)-Tetra-hydro-6-methyl-2H-pyran-2-yl]oxy]-9-ethyl-2,3,3a,5a,5b,6,9,10,11,12,13,14,16a,16b-tetrad-cahydro-14-methyl-1H-as-Indaceno[3,2-d]oxacyclododecin-7,15- dione 及Spinosyn D (Factor D CAS#131929-63-0) 或2-[(6-deoxy-2,3,4-tri-O-methyl-[alpha]-L-manno-pyranosyl) oxy]-13-[[5- (dimethyl-amino)-tetrahydro-6-methyl-2H-pyran-2-yl]oxy]-9-ethyl-2,3,3a,5a,5b,6, 9,10,11,12,13,14,16a,16b-tetradecahydro-4,14-methyl-1H-as-Indaceno[3,2-d]Oxacy-clododecin-7,15-dione组成的多杀菌素(Spinosad)残留许可限量:杏壳:19 ppm;坚果树14组:0.10 ppm;开心果:0.10 ppm;酸枣:0.10 ppm及石榴:0.30 ppm。 2009年9月24日,美国发布通报,美国环保署拟制定杀虫剂乙草胺的许可限量。 本最终法规规定了以下作物内/表杀虫剂乙草胺(Acetochlor) ,包括其代谢物及降级物的许可限量:轧棉副产品:4.0 ppm 未去纤维棉籽:0.6 ppm 大豆粉:1.2 ppm及大豆种:1.0 ppm。这些残留限量标准合格性只能通过检测乙草胺(acetochlor)、2-chloro-2''''-methyl-6-ethyl-N-ethoxymethylacetanilide及其含半EMA及半HEMA的代谢物来确定。(信息来源:中国技术性贸易措施网)
http://www.bioon.com/biology/UploadFiles/201202/2012021423545689.jpg约二千年以来,中医一直用一种通常称为常山的根提取物治疗疟疾,其中常山来自一类生长在西藏和尼泊尔的八仙花属植物。最近的研究表明,常山酮(溴氯哌喹酮)也可以用来治疗许多种自身免疫性疾病,其中常山酮是一种衍生自这种提取物活性成分的化合物。现在,哈佛大学牙科医学院的研究人员已经发现这种中草药提取物粉末背后的分子秘密。已证明常山酮(HF)触发一种阻断一类有害免疫细胞发展的应激反应通路,这类有害免疫细胞称为Th 17细胞,它被牵涉入许多自身免疫性疾病中。"HF防止自体免疫反应,不完全抑制免疫力",此项新研究的通讯作者、哈佛大学牙科医学院发育生物学教授 Malcolm Whitman说,"这种化合物能激发各种自身免疫性疾病的新治疗方法"。"这项研究是一个如何解决传统草药分子机制的令人兴奋的例子,它可导致生理调节的新见解和治疗疾病的新方法",Tracy Keller说,他是Whitman实验室的一名讲师,也是此文章的第一作者。这项研究包括一个马萨诸塞州总医院和其他地方的跨学科研究小组的研究人员,其中包括一个多学科小组的研究人员在和其他地方,它将被在线发表在2月12日的Nature Chemical Biology上。以前的研究已经表明,HF减少组织内疤痕、硬皮症(一种皮肤紧缩症)、多发性硬化症、瘢痕形成甚至癌演进。"我们认为,HF必须作用于有许多下游效应的信号通路", Keller说。在2009年,Keller和同事们报道,HF在不影响其他有益的免疫细胞情况下保护免受有害的Th17免疫细胞。自2006年被识别以来,Th 17细胞是暗含在许多象炎症性肠疾病、类风湿性关节炎、多发性硬化症和牛皮癣一样的自身免疫性疾病的"糟糕反应物"。研究人员发现,小剂量HF降低小鼠模型上的多发性硬化症。照这样,它是药物的新军械库其中之一,这种药物选择性地抑制自身免疫性病理不整体性地抑制免疫系统。进一步分析表明,HF某种程度上打开一个参与新近发现的称为氨基酸反应通路(AAR)的基因。科学家们最近才理解营养AAR传感通路在免疫调节与代谢信号中的作用。也有证据表明,它延长寿命并延迟热量限制动物研究中与年龄有关的炎症性疾病。作为一位分类自然资源保护学家,AAR让细胞知道它们什么时候需要保护资源。例如,当一个细胞感觉建造蛋白质的氨基酸的有限供应,AAR将阻断促进炎症的信号,因为发炎组织需要大量的蛋白质。"想想停电期间我们如何保养小果汁我们离开我们的设备上,上述聊天支持紧急呼叫",Whitman说,"细胞利用类似的逻辑"。对当前研究来说,研究人员调查研究HF如何激活AAR通路,寻找细胞用来将一个基因DNA编码翻译成构成蛋白质的氨基酸链的最基本过程。研究人员能追击称为脯氨酸的单一氨基酸,并发现HF靶向且抑制一种特异酶(tRNA合成酶,EPRS),这种特异酶负责将脯氨酸掺入通常含有脯氨酸的蛋白质中。当这发生时,AAR响应踢掉砸开并产生HF治疗的疗效。提供补充的脯氨酸逆转HF对Th 17细胞分化的效应,而添加回其他氨基酸没有逆转作用,为脯氨酸掺入建立HF特异性。补加的脯氨酸也逆转HF的其他治疗作用,抑制它抗疟疾的效应及与疤痕组织相关的特定细胞过程。再次,补充其他氨基酸却没有这样的效果。这样固定证据清楚地表明,HF特异地作用于受限制的脯氨酸。研究人员认为,HF治疗模仿了细胞内脯氨酸丧失,这激活AAR反应,并随后冲击免疫调节。研究人员还没完全了解氨基酸限制在疾病反应中的作用或限制脯氨酸为什么抑制Th 17细胞的产生。然而,"AAR通路显然是一个有趣的药物目标,且常山酮除了其潜在的治疗用途外是一种研究AAR通路的强大工具", Whitman说
麻烦问一下,有没有做5009.142的标准,吡氟禾草灵标准品怎么衍生,标准里面样品衍生了,没有说标准品的衍生过程
大家好,有没有检测吡氟禾草灵的,在检测的时候有那些经验技巧!谢谢!
哪位老师测过禾草灵和吡氟禾草灵,按照国标法GB23200.8和GB23200.121,都测不出来。有什么需要注意的吗?谢谢
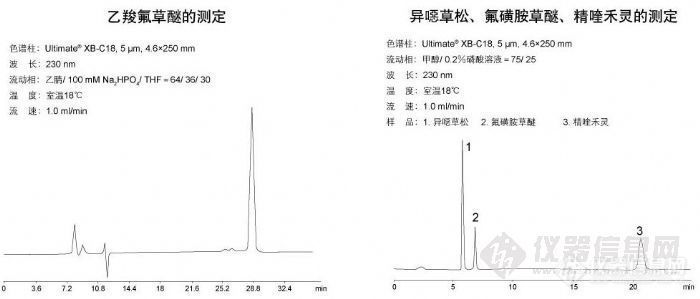
乙羧氟草醚的测定和异噁草松、氟磺胺草醚、精喹禾灵的测定http://ng1.17img.cn/bbsfiles/images/2009/11/200911131331_184260_1896702_3.jpg
求助:茶叶中除草剂吡氟酰草胺(DlFLUFENICAN)的检测方法。
请问谁做过吡氟禾草灵?5009.142的方法简直是太变态了,哪位大侠指点一下?
2011年6月14日,日本厚生劳动省发布G/SPS/N/JPN/278号通报:修订食品卫生法项下食品和食品添加剂标准和规范,涉及以下7种农兽药的最大残留限量:农药:1. 三氟羧草醚 Acifluorfen2. 二噻农 Dithianon3. 乳氟禾草灵 Lactofen4. 二甲戊乐灵 Pendimethalin5. 氟吡草胺 Picolinafen兽药:6. 左旋咪唑 Levamisole7. 甲苯咪唑 Mebendazole该通报的评议截止期为2011年8月8日,内容详见:http://members.wto.org/crnattachments/2011/sps/JPN/11_1819_00_e.pdf
有做氟磺胺草醚的么?想咨询下,扩项的加标水平如果加检出限,不能做出回收怎么办?
吡氟禾草灵相关的化合物一共有如下几种:1. Fluazifop2. Fluazifop-P3. Fluazifop-butyl4. Fluazifop-P-butyl5. Fluazifop-methyl其中1和2,3和4是同分异构体,3、4是1、2的丁基酯想请教大家,知道以上五种物质对应的中文名嘛?谢谢!实际检测过程中是检测其中哪几种物质?我看日本厚生省的关于吡氟禾草灵的检测方法分析的目标化合物是“吡氟禾草灵酸、吡氟禾草灵、精吡氟禾草灵酸、精吡氟禾草灵”四种物质,这四种物质分别对应的英文名称是?
同行们:好,最近有朋友做精吡氟禾草灵吗,为什么出峰时间和影响值都不稳定,其图如下:图1中:4.5-5min中和9min提取离子都是精吡氟禾草灵,图2在5--5.5min和8.5-9.5min都是精吡氟禾草灵图3:精吡氟禾草灵的峰有没有出峰有人做过这种物质吗,这种物质是由对映体,S和R,但精吡氟禾草灵应该都是R型,所以我觉得也不应该出现这种问题呀,不知道是不是水解的问题,因为这是一种酯类化合物,但我觉得是这个问题也不应该忽高忽低!下周再试一下!朋友们有什么高见,敬请发表意见!
高效氟吡甲禾灵(高效盖草能)标准,跪求标准,感谢各位大侠!
上世纪60年代。当时,杜邦公司开发出了首个脲嘧啶类除草剂—除草定,正式开启了该类除草剂研发的先河。而真正掀起脲嘧啶类除草剂开发热潮的是在上世纪90年代,当时人们对于该类除草剂的作用机理有了更深入的了解,发现脲嘧啶类除草剂属于原卟啉原氧化酶(PPO)抑制剂。杜邦公司在推出除草定后,又相继推出了异草定和特草定等产品。富美实的双苯嘧草酮以及先正达的氟丙嘧草酯均属于该类除草剂。而巴斯夫于2009年推出的苯嘧磺草胺(saflufenacil)更属于该类除草剂中的佼佼者。苯嘧磺草胺能够适用于多种生产系统和非耕地,在苗后或苗前均能使用;其次,适用作物多。苯嘧磺草胺能够用于包括谷物、玉米、棉花、水稻、高粱、大豆和果树等在内的30多种作物上;再次,防除谱广。苯嘧磺草胺能够防除90余种阔叶杂草,包括一些对三嗪类、草甘膦及乙酰乳酸合成酶抑制剂存在抗性的杂草。另外,它也具有作用快、残效期长等多种特性。http://ng1.17img.cn/bbsfiles/images/2017/02/201702010042_01_1623180_3.jpg2009年,苯嘧磺草胺在南美国家尼加拉瓜、智利和阿根廷三国登记。2010年,苯嘧磺草胺与精二甲吩草胺的复配制剂Verdict在美国获得登记,用于大豆。同年,苯嘧磺草胺正式登陆中国,以70%水分散粒剂(商品名:巴佰金)的形式面世,用于柑橘园和非耕地的杂草防除,由诺普信负责在中国市场的总经销。目前,苯嘧磺草胺已在美国、加拿大、中国、尼加拉瓜、智利、阿根廷、巴西和澳大利亚等国登记。苯嘧磺草胺可替代苯氧类除草剂2,4-D和磺酰脲类除草剂与草甘膦复配,可降低防治顽固性杂草对草甘膦的使用量。2014年,苯嘧磺草胺的全球销售额达到1.4亿美元。据巴斯夫公司预测,苯嘧磺草胺可实现3亿欧元的年峰值销售额。苯嘧磺草胺目前仍处于专利保护期中,其在中国的专利为巴斯夫于2001年申请的《尿嘧啶取代的苯基氨磺酰羧酰胺》,专利号为ZL01801896.3,对苯嘧磺草胺的化合物及合成方法进行了保护,该专利将于2021年4月30日到期.
[table=338][tr][td] [table=437][tr][td=1,1,157][size=16px][font=宋体]日文名称[/font][/size][/td][td=1,1,133][size=16px][font=宋体]英文名称[/font][/size][/td][td=1,1,147][size=16px][font=宋体]中文名称[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]EPN[/font][/size][/td][td][size=16px][font=宋体]EPN[/font][/size][/td][td][size=16px][font=宋体]苯硫磷[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]イミダクロプリド[/font][/size][/td][td][size=16px][font=宋体]IMIDACLOPRID[/font][/size][/td][td][size=16px][font=宋体]吡虫啉[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]オキサジクロメホン[/font][/size][/td][td][size=16px][font=宋体]OXAZICLOMEFONE[/font][/size][/td][td][size=16px][font=宋体]氯恶嗪草(去稗安)[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]ジクロシメット[/font][/size][/td][td][size=16px][font=宋体]DICLOCYMET[/font][/size][/td][td][size=16px][font=宋体]双氯氰菌胺[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]ノバルロン[/font][/size][/td][td][size=16px][font=宋体]NOVALURON[/font][/size][/td][td][size=16px][font=宋体]双苯氟脲[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]フェノキサニル[/font][/size][/td][td][size=16px][font=宋体]FENOXANIL[/font][/size][/td][td][size=16px][font=宋体]氰菌胺(禾草灵)[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]フェリムゾン[/font][/size][/td][td][size=16px][font=宋体]FERIMZONE[/font][/size][/td][td][size=16px][font=宋体]嘧菌腙[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]プレチラクロール[/font][/size][/td][td][size=16px][font=宋体]PRETILACHLOR[/font][/size][/td][td][size=16px][font=宋体]丙草胺[/font][/size][/td][/tr][tr][td][size=16px][font=宋体]ペンシクロン[/font][/size][/td][td][size=16px][font=宋体]PENCYCURON[/font][/size][/td][td][size=16px][font=宋体]戊菌隆[/font][/size][/td][/tr][/table][/td][/tr][/table]附件请参考。
大家使用GB 23200.108-2018 测植物源性食品中草铵膦残留量,多壁碳纳米管哪里采购的?[img]https://simg.instrument.com.cn/bbs/images/default/em09511.gif[/img]这个标准今天刚下载到,与大家分享。虽然之前有征求意见稿![img]https://simg.instrument.com.cn/bbs/images/default/em09502.gif[/img]
[color=#444444]乙羧氟草醚,需要检验其百分含量,请问各位大侠有具体的方法用[url=https://insevent.instrument.com.cn/t/Mp]气相色谱仪[/url]检测?据我所知,可以用有机液(比如甲醇)溶解,然后扣除溶解峰,就是主产品含量,请问大家,这个方法可行吗?甲醇检测方法和乙羧氟草醚检测的方法(温度控制方法初温,检测温度,)相同吗?[/color]
纳米中草药是指运用纳米技术制造的、粒径小于100nm的中药有效成分、有效部位、原药及其复方制剂,具有增加药物对血脑屏障或生物膜的穿透性等特点。纳米中草药技术(nanotechnology)是指在纳米尺度下对物质进行制备、研究和工业化,以及利用纳米尺度物质进行交叉研究和工业化的一门综合性的技术体系。纳米技术作为高新技术,可广泛应用于材料学、电子学、生物学、医药学、显微学等多个领域,并起着重要的作用。 一、纳米中草药的特点 1、原药纳米化后呈现新的药效或增强原有疗效中药被制成粒径0.1~100 nm大小,其物理、化学、生物学特性可能发生深刻的变化,使活性增强和产生新的药效。如灵芝通过纳米级处理,可将孢子破壁,并采用超临界流体萃取技术萃取出灵芝孢子的脂质活性物质,从而增强抗肿瘤的功效。 2、改善难溶性药物的口服吸收。在表面活性剂、水等存在下,直接将药物粉碎成纳米混悬剂,增加了药物溶解度,适于口服、注射等途径给药,以提高生物利用度。 3、增加药物对血脑屏障或生物膜的穿透性。纳米粒能够穿透大粒子难以进入的器官组织、血脑屏障及生物膜。如阿霉素α聚氰基丙烯酸正丁酯纳米粒(NADM)可以改变阿霉素的体内分布特征,对肝、脾表现出明显的靶向性,而血、心、肺、肾中的药物分布则减少。 4、靶向作用。在研究中发现,一味普通的中药牛黄,加工到纳米级水平后,其理化性质和疗效会发生惊人的变化,甚至可以治疗某些疑难杂症,并具有极强的靶向作用。 5、使药物达到缓释、控释。借助高分子纳米粒作载体等技术手段,可实现药物的缓释、控释。如雷公藤乙酸乙酯提取物固体纳米脂质粒有良好的缓释、控释功能。二、纳米中草药的制备技术及其进展 纳米中药的制备是研究纳米中药最基础的,也是最重要的问题。将纳米技术引入中药的研究,必须考虑中药组方的多样性、成分的复杂性,例如中药单味药可分为矿物质、植类药、动物药和菌物药等,中药的有效部位和有效成分又包括无机化合物和有机化合物、水溶性成分和脂溶性成分等,因此,针对不同的药物,在进行纳米化时必须采用不同的技术路线。此外,还必需考虑中药的剂型。纳米中药与中药新制剂关系十分密切,如何在中医理论的指导下进行纳米中药新制剂的研究,将中药制成高效、速效、长效、剂量小、低毒、服用方便的现代化制剂,也是进行中药纳米化所必须考虑的问题。纳米中药是针对中药的有效成分或有效部位进行纳米技术加工处理,开发中药的新功效。聚合物纳米粒可作为药物纳米粒子和药物纳米载体。药物纳米载体系指溶解或分散有药物的各种纳米粒,药物纳米载体包括纳米脂质体、固体脂质纳米粒以及纳米囊和纳米球。而对于不同类型的纳米中药,有不同的制备方法。三、纳米中草药的加工方法。 1、纳米超微化技术是改进某些药物的难溶性或保护某些药物的特殊活性,适用于不宜工业化提取的某些中药。如矿物药、贵重药、有毒中药、有效成分易受湿热破坏的药物、有效成分不明的药物。目前比较常用的是超微粉碎技术。所谓超微粉碎是指利用机械或流体动力的途径将物质颗粒粉碎至粒径小于10 μm的过程。根据破坏物质分子间内聚力的方式不同,目前的超微粉碎设备可分为机械粉碎机、气流粉碎机、超声波粉碎机。 2、机械粉碎法是利用机械力的作用来实现粉碎目的。边可君等采用自主开发的温度可控(-30~-50℃)的惰性气氛高能球磨装置系统制备纳米石决明。将石决明置于配有深冷外套的惰性气氛球磨罐中,同时装入磨球,磨球与石决明粉比保持在15:1~5:1范围,控制高能球磨机的转速(200~400 r/min)和时间(2~60 h),获得了平均粒度不大于100 nm的石决明粉末。 3、气流粉碎法是以压缩空气或过热蒸汽通过喷嘴产生的超音速高湍流气流作用为颗粒的载体。颗粒与颗粒之间或颗粒与固定板之间发生冲击性挤压、摩擦和剪切等作用,从而达到粉碎的目的。与普通机械冲击式超微粉碎机相比,气流粉碎产品粉碎更细,粒度分布范围更窄。同时气体在喷嘴处膨胀降温,粉碎过程中不会产生很大的热量。所以粉碎温升很低。这一特性对于低融点和热敏性物料的超微粉碎特别重要。 备注: 纳米中草药的常用方法较为普遍的有两种:一是喷雾干燥法,二是高能球磨法。 中药纳米超微化技术既丰富了传统的炮制方法,又能为中药的生产和应用带来新的活力。纳米产品目前已成为中药行业新的经济增长点。将这项技术应用于中药行业可以开发具有更好疗效、更优品种的纳米中药新产品。这将对中药行业的发展带来深远的理论和现实。
复方甘草口服溶液要避光保存,说是吗啡遇光易分解,但是在固定相萃取中并未避光,但是结果未产生太大影响,请文助威高手,复方甘草口服口服溶液是否对于避光不必太过在意
现有84.8%的乳氟禾草灵的药品,想要做薄层层析,但苦于没有找到合适的展开剂,请那位高手给指点一二,能提供三氟羧草醚的也可以,小弟不胜感激!
鼻血草别名:红活美化学成份:Melissa属植物的叶含鞣质约5%,又含挥发油。原形态:多年生草本,高0.6-1m,被短柔毛。具地下茎。茎四棱形,具分枝。叶对生,叶柄长2-25mm,密被短柔毛;叶片卵形,长1.2-6cm,宽9-30mm,先端急尖或短渐尖,基部圆形、钝或近心形,边缘具锯齿状圆齿,上面疏被短柔毛,下面靠近中脉两侧带紫色或伞部紫色,近无毛或仅沿脉被短柔毛。轮伞花序少花或多花,腋生;苞片小,具缘毛;花萼钟形,长6-8mm,外面被具节长柔毛,内面无毛、上唇3齿,齿短,下唇与上唇近等长,2齿,齿披针形;花冠白色或淡红色,长约1cm,外被短柔毛,内面无毛,上唇先端微缺,下唇3裂,中裂片较大;雄蕊4,前对较长,内蒙,花药2室;子房4裂,花柱略长于雄蕊,柱头2裂;花盘4裂。小坚果卵圆形。花期6-11月,果期7-11月。功能主治:清热,解毒。治风湿麻木,大麻风,吐血,鼻出血,皮肤瘙痒,疮疹,癫症,崩带。
请问除草剂密草通能溶于水吗?溶解度是多少?
苯醚甲环唑与草铵磷的液质联用检测GB/T20769-2008这个标准我怎么找不到这两种农药残留的检测方法啊,你们是怎么检测这两种农药的,母离子、子离子分别是多少啊
GB 2763-2012标准中有涉及草铵膦和丁醚脲两种农药组分,但值给出MRL和ADI值,无告知相应的检测方法可供参考。请各位色友帮忙哦。先谢了!
求吡喹酮、氯氰菊酯、扑草净、马拉硫磷、栀子、大黄、连翘、白芍、牡丹皮、人参、黄芪、甘草、黄连、黄芩、黄柏、木香等的液相色谱图!!!!!!!!!!!
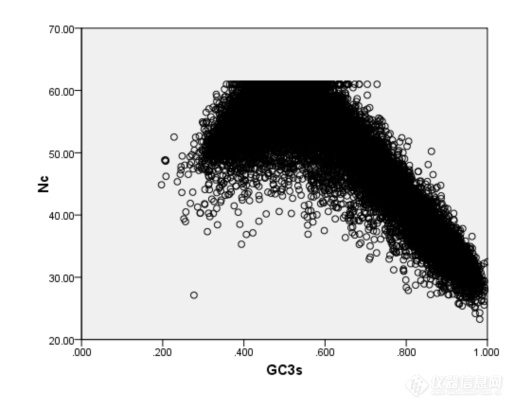
[align=center]短柄草全基因组密码子用法分析分析[/align]摘要:本研究运用CodonW程序分析了短柄草全基因组的密码子使用特性,并且通过对应分析探讨了若干重要因子对短柄草全基因组序列密码子用法的影响。结果表明短柄草基因组存在高[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量和低[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量的基因,它们在密码子使用上差异较大。Nc-plot曲线表明基因组的密码子组成受到碱基组成的影响;对应分析显示,在DNA水平上发生的核苷酸突变可能是造成短柄草基因组密码子使用偏好的主要因素;同时,基因长度和蛋白质疏水性对密码子的使用也存在一定偏性,但影响程度不大。确定了UUC等27个以G或C碱基结尾的密码子为“最优密码子”,研究结果可为短柄草基因的鉴定、表达、结构、功能等的深入研究提供参考。关键词:同义密码子偏好性,短柄草基因组,对应分析近年来,随着分子生物学的快速发展,许多小基因组的低等生物和高等模式生物的全基因组序列均被测定,为利用生物信息学方法挖掘海量基因组数据提供了便利。密码子是生物体内遗传信息传递的基本环节,是核酸携带信息和蛋白质携带信息间对应的基本规则。在长期进化过程中,任一物种的基因都会逐渐适应宿主的基因组环境,而形成特定的且符合宿主基因组的密码子用法,因此不同生物具有不同的密码子使用模式。以生物基因组数据为基础,研究其密码子使用模式,为深入研究基因的结构、功能和基因组进化,以及指导基因转化等具有重要意义。密码子具有简并性,生物在同义密码子的使用上并不是完全随机的,而是具有一定的偏向性,对有的密码子使用频率高,有的使用频率低,甚至避免使用,这种不均衡使用密码子的现象普遍存在于原核和真核生物中。早在20世纪70年代,人们在研究基因的异源表达时,就已经意识到密码子偏性的重要性[1],随着不同生物基因组数据的获得和各种数据库的构建,更多的研究者对密码子偏性的研究产生了浓厚的兴趣,尤其在分子进化,翻译调控等研究领域,通过对不同物种的密码子使用偏性的大量研究[2~4],发现不同物种的基因在密码子使用上存在着明显的偏性。 短柄草是一种广泛分布于温带地区的禾本科植物,与小麦,大麦和燕麦同属早熟禾亚科,原产于非洲北部,欧洲南部和亚洲中部,包含约10个亚种。该植物为一年生,自花授粉,植株高度15~20cm,生育期70~80d,柄草植株较小,适应性强,不象种植水稻那样需要严格的生长条件。生育期短,籽粒产量较高,一年可以繁殖4~5代,繁殖系数达140左右。未成熟胚和成熟胚愈伤组织诱导率高,农杆菌介导和基因枪介导的转化体系已经建立,胚性愈伤组织分化率90%以上,转化效率最高可达55%左右。基因组小,染色体少,DNA重复序列低,获得突变体容易,突变性状容易显现,具备了模式植物的所有基本特征。加之短柄草基因组序列与黑草麦,小麦,大麦等早熟禾亚科植物高度相似,很多重要农艺性状与温带禾草类植物相似,如株型,穗型,粒型,抗逆性,生长习性和病原菌等,其中麦类作物白粉病菌,条锈病菌和稻类作物瘟病菌都可侵染短柄草植株,引起相应症状[7]。其籽粒不含高分子量麦谷蛋白亚基,低分子量麦谷蛋白亚基也很少,并与小麦一样具有二倍体,四倍体和六倍体,因此短柄草是小麦等基因组庞大的重要农作物理想的模式植物,借此来获得目前小麦等早熟禾类植物中尚缺少的遗传信息和基因共线区,进而对小麦等重要植物进行基因定位,克隆,突变,测序和功能等方面的研究[8]。 目前,在短柄草的生物学、细胞学和遗传学特性方面开展了大量研究,并且其全基因组测序也基本完成[9],为深入研究其密码子用法提供了便利。因此本研究将以短柄草全基因组序列为基础,分析其基因的密码子用法特性和影响密码子使用的因素等,其研究结果将对指导转基因及对基因进行特定分子改造,提高其在短柄草中的表达效率和完善基因预测软件,提高基因预测和基因组注释准确性等均具有重要的参考价值,同时也为深入开展基因结构和功能,分子进化等研究提供理论基础。1.实验材料与方法1.1材料 短柄草全基因组DNA序列来源于短柄草官方数据库(http://www.brachypodium.org/node/8),根据基因组序列的注释信息,获得蛋白编码基因序列,为了减少长度较短的基因变异带来的样本误差,根据国际惯例,去除小于300bp的基因,去除中间不表达的密码子,终止密码子。编写程序提取剩下的蛋白编码基因的CDS(coding sequence)序列。1.2方法用codonw软件计算短柄草全基因组的密码子用法相关参数,主要包括有效密码子数(Effective Number of Codon,ENC)、基因的G+C含量([url=https://insevent.instrument.com.cn/t/Mp]gc[/url]%)、[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]3s%、相对同义密码子使用度(relative synonymous codon usage,RSCU)、氨基酸组分指数(平均亲水性值(gravy))、基因长度即氨基酸数(L_aa)。其中,有效密码子数(Effective Number of Codon,ENC)描述密码子使用偏离随机选择的程度,能反映密码子家族中同义密码子的非均衡性的偏好;其取值范围在20到61之间,即如果每种氨基酸只使用一种密码子则有效密码子数为20,如果各种同义密码子的使用机会完全均等,则有效密码子数为61,数值越小偏性越强。此值是以描述密码子使用偏离随机选择的程度,能反映密码子家族中同义密码子的非均衡性的偏好。基因密码子偏爱程度越大,ENC值越小。RSCU是指对于某种特定的密码子在编码对应氨基酸的同义密码子间的相对频率;[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]3s%表示同义密码子第三位碱基的G+C的含量。为进一步了解该家族基因密码子使用特征和影响密码子使用的因素,对7个基因的相对同义密码子使用度进行了对应性分析(correspondence of analysis,COA)。2 结果与分析2.1 基因的碱基组成对密码子使用的影响图一 短柄草基因NC值散点图[img=,515,409]https://ng1.17img.cn/bbsfiles/images/2019/10/201910311236371230_3093_3295053_3.png!w515x409.jpg[/img]2.2短柄草基因密码子使用特性的对应性分析[img=,690,535]https://ng1.17img.cn/bbsfiles/images/2019/10/201910311237226440_1452_3295053_3.png!w690x535.jpg[/img][img=,690,534]https://ng1.17img.cn/bbsfiles/images/2019/10/201910311237233450_935_3295053_3.png!w690x534.jpg[/img]2.3 确定最优密码子Phe UUU 0.05 (323) 1.23 (19733) Ser UCU 0.22 (990) 1.60 (23834) UUC* 1.95 (13527) 0.77 (12294) UCC* 2.55 (11715) 0.64 (9499) Leu UUA 0.02 ( 93) 0.83 (11755) UCA 0.14 (629) 1.52 (22651) UUG 0.16 (1003) 1.37 (19558) UCG* 1.53 (7023) 0.35 (5159) CUU 0.14 (847) 1.55 (21987) Pro CCU 0.22 (1306) 1.57 (17584) CUC* 3.38 (20676) 0.61 (8661) CCC* 1.35 (7940) 0.47 (5299) CUA 0.07 (452) 0.70 (9983) CCA 0.20 (1184) 1.62 (18078) CUG* 2.23 (13637) 0.94 (13401) CCG* 2.22 (13058) 0.34 (3792) Ile AUU 0.12 (398) 1.41 (21216) Thr ACU 0.10 (401) 1.46 (16515) AUC* 2.76 (9124) 0.70 (10557) ACC* 1.75 (7291) 0.66 (7397) AUA 0.12 (380) 0.89 (13461) ACA 0.12 (509) 1.56 (17636) Met AUG 1.00 (8512) 1.00 (20892) ACG* 2.03 (8478) 0.32 (3563) Val GUU 0.10 (693) 1.67 (23852) Ala [url=https://insevent.instrument.com.cn/t/Mp]gc[/url]U 0.14 (1914) 1.65 (26184) GUC* 1.71 (12491) 0.63 (9025) [url=https://insevent.instrument.com.cn/t/Mp]gc[/url]C* 1.98 (27398) 0.58 (9131) GUA 0.05 (349) 0.75 (10713) [url=https://insevent.instrument.com.cn/t/Mp]gc[/url]A 0.13 (1802) 1.48 (23459) GUG* 2.14 (15605) 0.95 (13562) [url=https://insevent.instrument.com.cn/t/Mp]gc[/url]G* 1.75 (24170) 0.29 (4678) Tyr UAU 0.05 (229) 1.28 (14480) Cys UGU 0.06 (194) 1.10 (9360) UAC* 1.95 (8126) 0.72 (8075) U[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]* 1.94 (6645) 0.90 (7595) TER UAA 0.42 (172) 0.82 (335) TER UGA 1.63 (665) 1.30 (530) UAG 0.94 (384) 0.87 (356) Trp UGG 1.00 (4992) 1.00 (10053) His CAU 0.15 (598) 1.42 (16785) Arg CGU 0.16 (750) 0.85 (6945) CAC* 1.85 (7568) 0.58 (6825) C[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]* 2.75 (12565) 0.49 (4043) Gln CAA 0.15 (627) 1.05 (20215) CGA 0.11 (500) 0.64 (5273) CAG* 1.85 (7975) 0.95 (18259) CGG* 1.92 (8761) 0.55 (4527) Asn AAU 0.12 (465) 1.31 (26650) Ser AGU 0.05 (235) 1.13 (16754) AAC* 1.88 (7141) 0.69 (13985) A[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]* 1.52 (7002) 0.77 (11441) Lys AAA 0.11 (552) 0.98 (27077) Arg AGA 0.10 (445) 1.94 (15854) AAG* 1.89 (9406) 1.02 (28423) AGG 0.96 (4387) 1.53 (12516) Asp GAU 0.15 (1344) 1.44 (39136) Gly GGU 0.11 (882) 1.34 (18423) GAC* 1.85 (16539) 0.56 (15322) G[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]* 2.53 (20795) 0.71 (9826) Glu GAA 0.17 (1437) 1.13 (36292) GGA 0.19 (1522) 1.26 (17423) GAG* 1.83 (15812) 0.87 (27746) GGG* 1.18 (9700) 0.69 (9476) 注:Number of codons in high bias dataset 372333 Number of codons in low bias dataset 915109标注*的密码子是(p 0.01)3 讨论密码子使用偏好是突变偏好、自然选择和遗传漂变等共同作用的结果,与碱基组成、翻译选择压力、基因表达水平、基因长度、蛋白质氨基酸组成、碱基突变频率和模式、mRNA二级结构稳定性等很多因素有关[17]。张晓峰[18]等研究表明,单子叶植物基因组的[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量在同义密码子使用偏性的产生过程中起着决定性的作用,同义密码子使用偏性强烈的基因往往偏爱使用C或G结尾的密码子,且第三位密码子突变往往是密码子偏好性发生变化的决定原因。短柄草基因密码子使用模式的调查表明其中有高含量的[url=https://insevent.instrument.com.cn/t/Mp]gc[/url],并且[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]3的含量高于[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]1和[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]2。这表明相对于以A和T结尾的密码子而言,这些基因偏好于使用以G或C结尾的密码子。从原核生物到真核生物的基因中,密码子使用偏好是一个被广泛研究的重要进化现象。研究发现,许多因素,比如碱基组成,基因表达水平,蛋白质疏水性等影响着密码子的使用。为了解释密码子使用偏好的起因,也有许多假设被提了出来。其中被广为接受理论是“选择——突变——漂移”模型。该模型认为在对偏好密码子的选择和通过突变-漂移对非偏好密码子的保留之间,同义密码子的使用偏性存在一种平衡。本文的研究结果显示,[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]3s值与ENC值密切相关,并且基因也位于第一轴线,揭示了碱基组成是影响短柄草基因组中的密码子使用偏好的主要因素。碱基组成是影响短柄草基因密码子使用的主要因素,基因长度和蛋白质的疏水性在短柄草基因密码子使用中也起到了一定的作用,相似的结果在水稻、小麦中被发现[15,19]。本研究发现,在基因长度和[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]之间存在很强的负相关性。这表明,高[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量的基因越短,密码子偏好就越大。可能的原因是富含AT基因的翻译效率比富含[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]基因的翻译效率更高,这种效率的差异对长的基因更为重要。通常,全基因组的基因表达值在许多多细胞真核生物中并不能得到,特别是基因表达水平在不同的组织和不同发育阶段不一样时。因此,要定量相当困难。在短柄草基因组中,目前还缺少相当数量的基因表达的准确数据。另外,我们发现[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量特别是在第三个碱基位置的[url=https://insevent.instrument.com.cn/t/Mp]gc[/url]含量较大的影响着密码子的偏好时,暗示着碱基突变可能是重要因素,同时,碱基突变又受控于翻译选择。所以,尽管基因表达水平影响着密码子的使用,但这影响还是远远小于核苷酸组成对密码子使用的影响。因此,我们没有进一步分析基因表达的影响。通过优化密码子,提高外源基因在微生物、植物、动物中的表达已有不少成功报道,而确定最优密码子可为合理有效进行密码子改造提供可靠信息。本文确定了UUC等27个密码子为短柄草全基因组的最优密码子。分析结果可为指导转基因及对基因进行特定分子改造,提高其在短柄草中的表达效率和完善基因预测软件,提高基因预测和基因组注释准确性等提供重要的参考价值。参考文献[1] Stanley D,Farnden K J F, MacRae E A. Plant a-amylases:Func-tions and roles in carbohydrate metabolism[J]. Biologia,Bratislava,2005.60(suppl l6):65-71[2] Smith AM. Zeeman SC, Smith S M. Starch degradation[J]. Annu Rev Plant Biol,2005,56(25):73-98[3] Asatsuma S, Sawada C, Itoh K et al. Involvement of α-amylase I-1 in starch degradation in rice chloroplasts[J]. Plant Cell Physiol,2005,4:858-869[4] Kaplan F, Guy C L. β-amylase induction and the protective role of maltose during temperature shock[J]. Plant Physiol, 2004, 1:1674-1684 [5] Kaplan F,Guy C L. RNA interference of Arabidopsis beta-amylase 8 prevents maitose accumulation upon cold shock and increases sensitivity of PSII photochem-ical efficiency to freezing stress[J]. Plant J.2005,44(13):730-743[6] Joho Mundy, Anders Brandt. Messenger RNAs from the Scutellum and Aleurone of Germinating Barley Encode (lm3,14)--D-Glucanase, a-Amylase and Carboxypeptidase[J]. Plant Physiol, 1985,79(5):867-871 [7] 言普,李桂双.高压对水稻种子细胞膜透性和淀粉酶活性的影响[J]. 浙江大学学报(农业与生命科学版),2007,33(5):174-179[8] Monica M, Sanwo and Darleen A. DeMason. Characteristics of a-Amylase during Germination of Two High-Sugar Sweet Corn Cultivars of Zea mays L[J]. Plant Physiol, 1992,99(8):1184-1192[9] Goldman N , Yang Z. A codon based model of nucleotide substitution for protein coding DNA sequences[J]. Molecular Biology and Evolution,1994,11(9):725-736[10] Schmidt W. Phylogeny reconstruction for protein sequences based on amino acid properties[J]. Mol Evol,1995,41(8) :522-530[11] 时成波, 吕安国.改造稀有密码子提高SEA蛋白表达量[J]. 生物工程学报,2002,18(4):477-480[12] Ghosh T C , Gupta S K, Majumdar S. Studies on codon usage in Entamoeba histolytica[J]. Int J Parasitol,2000,30(6): 715-722[13] Musto H, Cruveiller S. Translational selection on codon usage in Xenopus laevis[J].Molecular Biology and Evolution,2001,18(9):1703-1707[14] 廖登群,张洪亮等. 水稻(Oryza sativa L.)a-淀粉酶基因的进化及组织表达模式[J]. 中国农业大学学报,2009,14(5):1-11[15]刘汉梅,何瑞. 玉米密码子用法分析[J]. 核农学报,2008,22(2):141-147[16] Jia M, Luo L. The relation between Mrna folding and protein structure[J]. Biophys Res Commum, 2006,343(4):177-182[17] 赵耀,刘汉梅. 玉米waxy基因密码子偏好性分析[J]. 玉米科学,2008,16(2):16-21 [18] Wang H C,Hickey D A. Rapid divergence of codon usage patterns within the rice genome[J].BMC Evol Biol,2007,15(8):347-356
求大豆油品中以下农残的国标方法:1. 倍硫磷2. 腐霉利、氯氟氰菊酯、高效氯氟烃菊酯、氯氰菊酯、高效氯氰菊酯、多效唑3. 氟吡甲禾灵、高效氟吡甲禾灵、噻虫啉4. 敌草快我现在改成悬赏去
求助乙氧氟草醚检测,是主含量检测不是残留,谢谢







 我要推广仪器
我要推广仪器
 下载APP
下载APP