农膜硫氯检测有相关标准吗?元素分析仪做出的结果和ICP的结果不一致
[font=仿宋_GB2312][size=21px][color=#6b6b6b]废弃农膜会破坏耕作层的土壤结构,使土壤空隙减少,降低土壤的通气性和透水性,使微生物和土壤动物的活力受到限制,同时不利于水分和营养物质在土壤中的传输,影响农作物对水分和营养物质的吸收,阻碍了农作物种子发芽、出苗和根系生长,造成农作物减产。[/color][/size][/font]
据国外媒体3月27日报道,提起机器,许多人可能马上会联想起那些钢铁铸造的大家伙,伴随着庞大的体积与轰鸣的马达声。但是,在微观世界里,还有一种科学家正潜心研制的超微小机器--分子机器。它们的尺度只有纳米(十亿分之一米)大小,但是却有望为人类带来许多福祉。日本科学家创造出了世界上最小的剪刀——分子剪刀,剪刀通过光就能启合。研究人员表示,这种新颖的剪刀可能有助于调控基因、蛋白质和人体内的其他分子。剪刀仅3纳米或十亿分之一米长,比紫色光的波长还要小100倍。这种分子装置是由东京大学的研究人员设计而成的,就像真正的剪刀一样,它也是由枢轴、手柄和刀片组成。研究人员将他们的发明公布于25日在芝加哥举办的美国化学学会年会上。 刀片由碳、氢构成的苯基环制成,枢轴是一种叫做手征性二茂铁的分子,它的结构基本上像一个三明治,是一个圆形铁原子处在两个碳原子板之间。碳原子板能围绕铁原子自由旋转。手柄是亚苯基团的有机化学结构。它与一种对光有反应的分子——偶氮苯连在一起。当可见光照在此剪刀上后,偶氮苯就会张开,从而让把柄打开,刀片合上。但是,当紫外线光照射剪刀时,会产生相反的效果。 研究人员声称,他们的剪刀可以像钳子一样牢牢地抓住分子,操控分子,也就是说能来回扭曲分子。研究人员阿伊达说:"这是首个分子机器能通过光来操作其他分子,这种工作的原理对于未来分子机器人的发展有着重要的作用。" 研究人员目前还在开发能够遥控操作的一把更大的分子剪刀。东京大学的一位研究人员Kazushi Kinbara说:"此剪刀能用于身体内部,可以通过近红外线操作,能进入人体更深的部分。" [color=red]【由于该附件或图片违规,已被版主删除】[/color]
MicroRNAs (miRNAs) 是真核生物中广泛存在的一种长约21到23个核苷酸的RNA分子,可调节其他基因的表达。miRNA来自一些从DNA转录而来,但无法进一步转译成蛋白质的RNA(属于非编码RNA)。这些RNA从初级转录本(primary transcript),也就是pri-miRNA,转变成为称为miRNA前体(pre-miRNA)的茎环结构,最后成为了具有功能的miRNA。细胞借助于一种特殊的机器——“微处理器”复合物("Microprocessor" complex)将这些长链的miRNA前体切断,然后将它们转变为较短的功能性的miRNA形式。生成的miRNAs与特异性信使RNA(mRNA)分子结合,抑制转录后基因表达,在调控基因表达、细胞周期、生物体发育时序等方面起实验室耗材重要作用。在动物中,miRNAs通常可以调控成百上千不同基因的表达。然而微处理器复合物也可以切断其他类型的RNA,例如mRNAs,这些mRNAs有时会生成与miRNAs靶位点相似的不稳定结构。裂解这种错误的RNAs有可能对生物体带来灾难性后果。在近期发布在《自然结构与分子生物学》(Nature Structural and Molecular Biology)杂志的一篇论文中,来自魏茨曼科学研究所分子遗传学系的Eran Hornstein博士、Naama Barkai教授和前博士生Omer Barad与Mati Mann侧重了解了这种微处理器在miRNAs生成中平衡有效性和特异性之间相互作用的机制。“一方面,它不应该过于有效,因为这可能会导致裂解不必要的非特异性RNA底物为代价。另一方面,也不应该太‘严格’因为过度的特异性会带来不充分加工真正miRNAs的风险,”Hornstein说。在一个跨学科研究项目中,科学家们利用数学建模确定了微处理器系统的特征,然后在细胞中测试了它们的理论。他们预测有效性和特异性之间的平衡是通过一个反馈回路维持,在这个人血清 混合反馈回路中微处理器检测到细胞内可用pre-miRNA的数量,相应地改变自身的生产。在小鼠和人类组织中检测这一假说,研究人员能够证实微处理器确实调控了miRNA前体的水平,当细胞充满miRNA前体时则提高自身的生产,或在前体量下降时作出反应停止生产。这是通过消化一种在结构上与miRNA相似的Dgcr8 mRNA而实现的。通过与miRNAs维持水平一致,微处理器由此减少了切断靶RNAs的机会。由于合成小RNAs是对于多种疾病一种可能的新疗法,这一研究或可直接促成未来有效的治疗。此外,许多其他的生物系统需要兼顾有效性和特异性,该研究小组的结果表明许多可能是细胞培养以相似的方式来实现的。
时下,物价暴涨,到底是什么原因,大家众说风云。食品、蔬菜、农产品、服装等均在提价。国家政府也在某些领域开始实施调控干预价格,这种调控是否长期有效,还是一时的打击?一段时间后,这种暴涨是否卷土重来?真的不得而知!
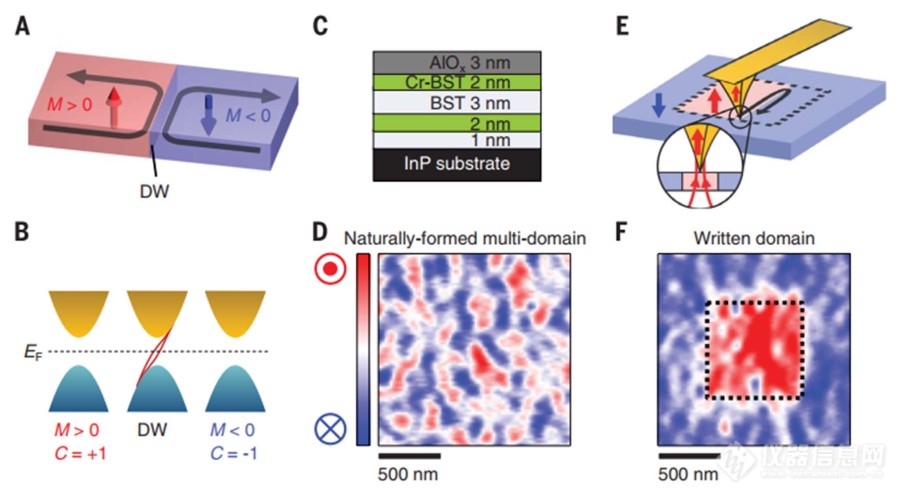
拓扑绝缘体,顾名思义是绝缘的,有趣的是在它的边界或表面总是存在导电的边缘态,这是拓扑绝缘体的独特性质。近期,理论预测存在的拓扑绝缘体在实验上被证实存在于二维与三维材料中,引起了科研界的大量关注。通常二维电子气体系中存在着量子霍尔效应,实验中观测到了手性边界态存在于材料的边界。在三维体材料的拓扑绝缘体中实验上可观测到反常量子霍尔效应。 K. Yasuda, Y. Tokura等人利用德国attocube公司的低温强磁场磁力显微镜attoMFM在0.5K温度与0.015T磁场环境下,证实了拓扑绝缘体磁畴壁的手性边界态的可调控性能,不同于之前实验上观测到的拓扑绝缘体中自然形成随机分布的磁畴中的手性边界态。Y. Tokura等人基于Cr-掺杂 (Bi1-ySby)2Te3制备了拓扑绝缘体薄膜,基底是InP(如图1C)。图1D为在0.5K极低温下使用MFM测量的材料中的磁畴分布,可以清晰看到自然形成的随机分布的大小与形貌不一的磁畴。通过使用MFM磁性探针的针尖在0.015T的磁场环境下扫描样品区域成功实现了对材料磁畴的调控。图1F为调控后样品的磁畴情况,被探针扫描过的区域,磁畴方向保持一致。[align=center][img=,500,273]http://ng1.17img.cn/bbsfiles/images/2018/07/201807311331396935_7457_981_3.jpg!w690x378.jpg[/img][/align][align=center]图1: A&B 拓扑绝缘体磁畴调控示意图;C 拓扑绝缘体材料结构;D attoMFM实验观测自然形成多个磁畴; E&F MFM探针调控磁畴[/align][align=center][/align][align=center] 该拓扑绝缘体磁畴反转的性能随磁场大小变化的结果也被仔细研究。通过缓慢改变磁场,不同磁场下拓扑绝缘体样品的磁畴方向可清楚地被证实发生了反转(见图2)。通过观察,随机分布气泡状磁畴(0.06T磁场附近)一般的大小在200纳米左右。[/align][align=center][/align][align=center][img=,500,206]http://ng1.17img.cn/bbsfiles/images/2018/07/201807311339098931_5066_981_3.jpg!w690x285.jpg[/img][/align][align=center]图2: A 霍尔器件电测量结果;B attoMFM观测不同磁场下拓扑绝缘体的磁畴情况[/align][align=center][/align][align=center] 不仅通过attoMFM直观观测分析磁畴手性边界态调控,电学输运结果也证实手性边界态的调控。图3为在温度0.5K的时候,拓扑绝缘体电学器件以及相应的电学测量数据。数据表明,霍尔电阻可被调控为是正负h/e2的数值,证实了不同磁畴的手性边界态的调控被实现。作者预见,该实验结果对于低消耗功率自旋电子器件的研究提供了一种可能的途径。[/align][align=center][/align][align=center][img=,500,565]http://ng1.17img.cn/bbsfiles/images/2018/07/201807311333567372_456_981_3.jpg!w690x780.jpg[/img][/align][align=center]图3:拓扑绝缘体制备器件反常量子霍尔效应结果证实磁畴手性边界态调控[/align][align=center][/align][align=center][img=,500,303]http://ng1.17img.cn/bbsfiles/images/2018/07/201807311334450730_967_981_3.jpg!w690x419.jpg[/img][/align][align=center]图4:拓扑绝缘体磁畴手性边界态调控相关设备—低温强磁场原子力磁力显微镜[/align][align=center][/align][align=center][/align]低温强磁场原子力磁力显微镜attoAFM/MFM主要技术特点:-温度范围:mK...300 K-磁场范围:0...12T (取决于磁体)-样品定位范围:5×5×5 mm3-扫描范围: 50×50 μ㎡@300 K, 30×30μ㎡@4 K-商业化探针-可升级PFM, ct-AFM, SHPM, CFM等功能参考文献:“Quantized chiral edge conduction on domain walls of a magnetic topological insulator” K. Yasuda, Y. Tokura et al, Science 358, 1311-1314 (2017)
霍华休斯医学研究所,Baylor医学研究所的科学家们近期在PloS One上发表最新研究性文章,文章标题为:Big Genomes Facilitate the Comparative Identification of Regulatory Elements,该文章解析了基因组大小对基因组学的研究带来的影响。基因组越大则更容易找出控制基因活性的DNA区域。在小基因组上,功能性元件紧紧地结合在一起。而在大基因组上,功能性元件分得比较散,于是也更容易找到控制基因活性的区域。 基因组分为结构基因和调控基因,要从基因组上找到功能元件并不难,难的是找到调控基因表达的机制,因此,对小的基因组来说,紧凑的结构给寻找调控区域带领更多的困难,而相对来说大基因组却容易多了。功能元件散落在基因组上,更便于寻找调控区域。大的基因组更便于研究非编码DNA和RNA,对研究基因调控也更为有利。而目前,研究生命的遗传物质DNA的科学家一直觉得,基因组越小越受欢迎,因为操作简单,可以节省大量的时间和精力,尤其在金钱方面也能更节约成本,测序的费用更低。甚至有科学家说,基因组小则基因排列更紧凑,垃圾DNA也越少。 [img]http://www.instrument.com.cn/bbs/images/affix.gif[/img][url=http://www.instrument.com.cn/bbs/download.asp?ID=137848]Big Genomes Facilitate the Comparative Identification of Regulatory Elements[/url]
[font=微软雅黑][size=16px][color=#161616]属于农业固体废物,不属于危险废物。根据《工业固体废物名录》,废塑料属于第13类固体废物;在《中华人民共和国固体废物污染环境防治法》第20条,明确表示畜禽粪便属于固体废物范畴。[/color][/size][/font]
请问那里有可以调控温度的 减压干燥仪?
版友们的仪器室,夜间有开空调控温吗,请说明相关理由。
[color=#333333]为抑制房价出现过快上涨苗头,近期各地陆续加大房地产调控力度,不断出台新政策、新措施。同时,住房租赁市场建设稳步推进。一系列举措表明地方政府坚持调控的信心和决心不会动摇,“房子是用来住的,不是用来炒的”这一定位不会改变!为什么国家越调控,房价越长的厉害呢?[/color]
由中国农业科学院北京畜牧兽医研究所王加启研究员主持完成的国家“十五”奶业科技重大专项“奶牛共轭亚油酸(CLA)合成调控机理研究及其产品开发” 研究成果已于近日荣获北京市科学技术一等奖。 该项成果以建立CLA定量检测技术、实时定量PCR技术等支撑技术为手段,以系统研究瘤胃微生物氢化规律和乳腺去饱和酶合成CLA的机理为理论探索的核心,以建立日粮调控、瘤胃发酵调控和乳腺合成调控有机结合的CLA原料奶生产技术体系为目标,同时开展CLA牛奶加工特性和免疫功能的延伸研究,最终开发出CLA牛奶产品,并在研究推广规范化饲养管理技术体系的基础上建立起优质功能牛奶生产基地。 该项成果在奶牛CLA合成机理、CLA牛奶生产技术体系等方面取得了重大突破和理论创新,在国内外率先开发出CLA液态奶产品,整体研究处于国际领先水平。在调控理论方面,明确提出瘤胃调控的重点是增加trans-11油酸的累积量,阐明日粮亚油酸和鱼油源性脂肪酸对乳腺CLA合成关键酶SCD的mRNA表达影响较小,主要通过增加前体物产量对奶牛CLA合成发挥底物效应;在调控技术方面,通过日粮组合、瘤胃发酵和奶牛个体筛选等单项技术的集成,将牛奶的CLA含量从10mg/100ml左右提高到40mg-90mg/100ml;在产品开发方面,研究建立CLA牛奶生产加工规范和产品质量标准体系,并开发出CLA纯牛奶产品。CLA牛奶与普通牛奶相比,饱和脂肪酸的比例从70%降低到61%;高胆固醇源性脂肪酸从45%降低到36%;在总CLA中,c9t11CLA的含量达85.6%,具有纯度高和未知因素少的优点。 该项成果建立的奶牛规范化饲养管理技术体系从2003年开始在全国2万头奶牛中推广,累计使养殖户增加纯收益3400万元,从2004年开始选择部分牛场作为CLA牛奶生产基地,到2005年底已经有3100吨CLA纯牛奶产品销售,平均每吨增加收益3000元以上。预计经过5年推广,有望培养出1000万以上的稳定消费人群,形成年产值5亿元以上的优质功能牛奶产业,给农民增加1亿元以上的收入。
科技日报10月30日报道 谷粒大小是决定水稻产量及品质的重要因素之一。最近,华中农业大学张启发院士课题组在谷粒大小和粒型的调控研究方面取得重大进展。本周出版的《美国科学院院刊》(PNAS)发表了该组研究论文,证实水稻中GS3基因控制水稻籽粒大小,发现了该基因中控制籽粒大小的关键区域OSR(OrganSizeRegulation)。 据文章第一作者、生科院博士生茆海亮介绍,张启发课题组从1997年开始对控制水稻粒形基因GS3研究,直到2006年找到这个基因,历时9年。在此基础之上,该组进一步证实GS3是调控谷粒大小的主要基因,揭示了基因所编码蛋白的结构与功能之间的关系。他们发现该基因编码的蛋白存在相互对抗的前后两个部分,其中前一段(N-端)是控制粒形的关键区域,即OSR,后一段(C-端)对OSR的功能有抑制作用,GS3蛋白内首尾两部分之间的“博弈”最终决定籽粒的大小。研究结果表明,没有GS3蛋白的品种的稻谷为长粒型(长度约10毫米),含有完整GS3蛋白的水稻品种粒型中等(约8毫米),而只有OSR的水稻品种的谷粒为短粒(约6毫米)。 研究还发现,几乎所有优良粳稻品种都带有完整的GS3蛋白,表现为中等粒型,优良长粒型籼稻品种的GS3蛋白无功能,通过对该基因的导入和替换,能有效地改变水稻品种的粒型。 茆海亮还提到,他们在玉米、大麦、大豆等其他物种中也发现了GS3同源的基因,并且OSR在这些同源基因中都存在,说明这些基因有可能也控制着相应物种种子大小。因此该研究成果在品种改良中将有着重大应用前景。该基因变异可作为分子标记直接应用于水稻籽粒大小的选育,提高水稻的产量。另外,根据水稻的研究信息,可以对其他物种的GS3同源基因进行克隆,从而指导相应物种的品种改良。
近日,国际著名学术期刊《血液》在线发表了中国科学院上海生命科学研究院/上海交通大学医学院健康科学研究所分子风湿病学研究组的最新研究发现:来自于同一前体的miR-155和miR-155*协同调控浆细胞样树突状细胞(pDC)I型干扰素的产生。 系统性红斑狼疮是一种以T细胞功能缺陷和B细胞过度活化及多种自身抗体产生为特点的自身免疫性疾病。已有的研究表明,I型干扰素的过度产生在系统性红斑狼疮的发病过程中起着十分重要的作用。它通过直接作用于T细胞和B细胞,促进自身免疫反应。因此,如何能够有效调控I型干扰素的产生,对于该自身免疫疾病的治疗有十分重要的意义。 近年来,microRNA作为一种非编码RNA分子,被证明在免疫调节的各个方面均有很重要的作用。因此,在沈南教授的指导下,整合中国科学院上海生命科学研究院健康科学研究所的基础研究力量和上海交通大学附属仁济医院风湿科的临床优势,周海波和黄新芳等深入研究了microRNA 在I型干扰素主要产生细胞——浆细胞样树突状细胞(pDC)中的调控作用。 该研究发现,pDC激活后伴随着大量I型干扰素的产生,miR-155*和miR-155分别在不同的时间被显著诱导。MiR-155*主要在早期被诱导,而miR-155则主要在刺激的后期被诱导。进一步研究表明,miR-155*通过靶向IRAKM,促进I型干扰素的产生,而miR-155通过靶向TAB2,抑制I型干扰素的产生。这些结果表明,它们在pDC活化的不同阶段协同发挥作用。此外,通过对miR-155和miR-155*产生机制的研究,发现pDC自身分泌的I型干扰素以及被激活的KHSRP蛋白可以在转录后水平反向调控miR-155和miR-155*的产生,这一结果解释了来自于同一前体的miR-155*和miR-155却能在不同的时间点被诱导的原因。 该研究不仅揭示了来自于同一前体microRNA和microRNA*的产生,在同一刺激过程中可以被精确调控,从而使它们能够协同调控这一过程,而且阐明了新的有效调控I型干扰素产生的机制,为系统性红斑狼疮疾病的的治疗提供了新的理论依据和潜在靶点。 该项工作得到国家科技部、国家自然科学基金和上海市科委的经费支持。
Cell文献:研究发现新的免疫系统调控分子开关
http://www.bioon.com/biology/UploadFiles/201112/2011123010390194.jpg 图片来源:“我的健康日报”网站近日,国际著名杂志International Journal of Cosmetic Science刊登了国外研究人员的最新研究成果“Population genetic segmentation of MHC-correlated perfume preferences。”,文章中,作者揭示了女性对香水味道喜好是受基因调控的;香水是每个女人最爱的产品,瑞士一项研究发现,女性对于香水味道的喜好,都会受到一组名为MHC的基因调控,该基因也决定了每个人会受到哪一种异性的气味所吸引。据报道,瑞士联邦技术研究所日前对116名受访者进行了不同气味的测试,包括肉桂、玫瑰、雪松等10种物质,而每次的测试浓度与环境都不相同。结果发现,最受欢迎的气味是“香树脂”,而“香根草”因为气味较平淡,最不受欢迎。研究指出,在不同气味的刺激下,人类受基因MHC控制会对味道呈现出强度不同的反应。英国剑桥大学专家莱斯利纳普表示,同一款香水给不同的人使用,会产生不一样的反应,而MHC基因将来也会被扩大研究。另一名瑞士专家德金德也表示,MHC基因与异性气味的关系是在女性闻嗅男性衬衫味道的实验中发现的,他还指出,90年代初期就曾发现过人体对于气味的喜好与免疫基因有关。(
[size=4][font=黑体]简介:量子调控研究是国家中长期科技发展战略规划的重要内容。近日,中科院物理所纳米物理与器件实验室高鸿钧研究组与谢心澄研究员及英国利物浦大学Werner A. Hofer教授合作在单分子自旋态的量子调控研究中取得新进展[/font][/size]量子调控研究是国家中长期科技发展战略规划的重要内容。近日,中科院物理所纳米物理与器件实验室高鸿钧研究组与谢心澄研究员及英国利物浦大学Werner A. Hofer教授合作在单分子自旋态的量子调控研究中取得新进展。他们发现在酞菁铁分子Kondo效应中由于分子中心铁原子在金属表面的吸附位置不同对Kondo效应产生很大影响。相关研究结果发表在9月7日出版的《物理评论快报》(Phys. Rev. Lett. 99, 106402 (2007))上。这是首次报道吸附位置对单分子Kondo效应的调控作用,为单分子自旋态的量子调控及其在量子信息中应用研究提供了新思路。 Kondo效应是指磁性杂质中的局域自旋与自由电子强关联相互作用所引起的一系列低温反常现象。近年来,扫描隧道显微镜技术的迅速发展使人们能够精确地测量单个磁性原子或分子在金属表面上的Kondo效应,而在原子尺度上探索影响Kondo效应的因素是实现单分子自旋态量子调控的关键。 物理所高鸿钧研究组利用低温扫描隧道显微镜及扫描隧道谱,在对吸附在金表面的磁性分子酞菁铁的测量中,发现了Kondo温度高于室温的Kondo效应,并发现分子中心铁原子在金表面的吸附位置对Kondo效应影响很大。他们发现酞菁铁分子在金表面存在两种吸附取向,虽然在分子中心测量的扫描隧道谱显示两种分子取向都存在Kondo效应,但是彼此却存在很大差别。这种差别主要表现在两个方面:根据Fano理论拟合的Kondo温度,以及扫描隧道谱在费米面附近的线型。第一性原理计算及实验测量表明,两种取向的分子的中心铁原子吸附在金表面的不同位置:第一种分子取向,铁原子吸附在金表面两金原子之间的桥位置;第二种分子取向,铁原子吸附在金表面金原子的正上方。他们的理论分析表明,分子中心铁原子在金表面的吸附位置不仅影响到局域自旋与自由电子耦合相互作用的强弱,而且还会影响扫描隧道谱测量中隧穿电子的通道。 近年来,高鸿钧领导的研究组对纳米功能结构材料的调控生长、机制与物性等进行了系列研究(如:Phys. Rev. Lett. 97, 246101 (2006);97, 156105 (2006);96, 226101 (2006);96, 156102 (2006);Adv. Func. Mater. 17, 770 (2007))。根据该工作观察到的吸附位置对单分子Kondo效应,他们提出了调控单分子自旋量子态的可能途径:1)通过基底上不同位置或不同基底的物理化学性质(如:Phys. Rev. Lett. 97, 156105 (2006));2)通过调节纳米分子体系中非功能性侧链(如:Phys. Rev. Lett. 96, 226101 (2006))。这对量子调控和量子信息研究具有重要意义。 以上工作得到了国家自然科学基金委、国家科技部和中国科学院的资助。
原帖地址:http://ehome.online.sh.cn/main.php?fid=721&cid=22&sf=3&topicid=1216675调控房价,用增加成本的方式,房价是决无降价的道理的:土地、税费、利率、配套、城建、建材、人工、营销、公关、利润。。。。。。都上涨,却要控制房价,岂不自欺欺人。只不过这些成本升跌,大家还看得见,还能理解,“肉烂在锅里”,毕竟有帐可查、有据可凭。 权利、利益则是无形的手、一幅巨网,被掏空腰包的,往往还糊里糊涂。计划经济遗留下来的政企不分,钱权合一,一方面造成政府机构间的权利相争、利益博弈,另一方面各自为政、诚信丧尽。 权力部门调控房价?有几个真正是出于维护老百姓利益而关注房价涨、跌的? 发改委感兴趣的是房地产转制问题;国土局关心的是集体土地有多少变国有;银行担心的是存、贷利益;建设部操心的是多少部建房可转商品房;国税局只怕收不到税;财政部为赤字睡不好觉;地方政府要的是政绩工程;开发商惦记的是利润;房地产交易中心希望交易越火越好;老百姓盼的是。。。。。。 现今中国,没有一个行业象房地产所面临那么多的困惑和尴尬: 房产交易不仅要交契税、印花税,还要交营业税、土地增值税、所得税。。。。。。俨然就是一种投资行业,但真要说买房是正当投资、鼓励投资房地产,招来的一定是比侵权盗版、发廊小姐更多的非议和指责。 如果说房地产是商品、消费品,在中国所有的商品中,没有一种商品在使用、转让中还要交纳那么多的税、费。 投资受“公司法”、企业法、“合同法”、工商管理法规等保护;商品(消费品)受“消保法”保护,房地产受什么保护呢?说它是投资,却处处受限制、随时被“调控”;说它是商品,土地和房产的出让方----国家,却可以不顾“购房交易合同”、“房地产产权证”和已纳税的约束,土地、房屋随时可能被征用、动迁;权力部门以调控为由,单方面更改责、权、利,侵害“原有房”和“已购房”业主权益,岂不是违约、“霸王条款”行为?让人翘首以待的一部“物权法”,却阻力重重,迟迟出不了台。 出租车、地铁、电信业、水电。。。。。。调价都要实行听政制;银行跨行查询收费,人大代表激呼反对;路边的排档城管会砸;随便停车交警要罚;投诉家具、家电质量差,有消费者协会撑腰。。。。。。而房地产业主权益受到侵害,动辙数万,怎么却没人管没人问呢?原因很简单:老百姓、企业好管;权力机关、上级部门,谁敢管?! 股市作为资本投资市场,直到今天既不收营业税,也不收所得税,原因据说是因为股票有涨有跌,涨了收所得税、跌了呢?那么作为比股市起步晚的房地产同样也有涨有跌,现在,涨了要收所得税,跌了是不是要退税、补贴呢? 房地产本来就流转性差,买卖过程中已经收取了契税(即交易税),等同于已经收了营业税。房地产所得主要源于土地增值,既然征了土地增值税(有的已经开征,有的将包含在物业税中)再征所得税,征了契税再征收营业税,是不是有重复征税之嫌?如果说土地增值税和所得税,契税和交易税或营业税不是一个税种,那么其它行业、其它商品买卖为什么收了营业税就不征契税,企业收了增值税就不再征营业税呢? 看了国税局的征收房地产交易个人所得税通知,至少有两点不解,在所有房屋原值计算中,为什么唯有房改房(北京称‘部建房’)的原值是含糊其词的呢?如果按原来真正支付的购房款为依据,那么部建房所得是多少呢?该如何交税呢?更不要说还有货币分房、房贴呢。再说什么人才有“超标准”的部建房?按同一地区的经济适用房计算原值?原来的房改房或部建房周边有经济适用房吗?不就是想怎么定就怎么定吗?就如货币分房,大家都是90平米,似乎很公平,其实老百姓每平米是按3000元计算,而当头的按多少算,谁也不清楚,一万、二万还是更多?记得多年前,笔者单位的领导花了一百多万,为自己买了一套当时最豪华的外销公寓,有人举报到部里,纪委下来检查,结论是:虽然房子好了点,但并没错误,按其级别还没达到面积标准,以后货币分房时再补差。要知道当年一套同等面积的普通商品房才一、二十万,还有那么些头发花白的老知识分子等着分房,当然最后还是有几句安慰话的:大家的困难领导是知道的,但困难总要一步步解决吗,总要有先有后吗,要有耐心,要理解领导的难处、苦处。 记得曾有报道,华北某油田为了解决下岗职工再就业,上岗条件是必须要有“离婚证”,一时间‘突击离婚’搞得民政处门庭若市。现在,规定自住五年以上,家庭唯一的住房,可以免征所得税。“家庭唯一”包不包括“离婚单身”?看来许多人卖房前,要么先“分家”去派出所,要么先“离婚”去民政局,这下婚姻登记处该热闹了,就业笑话眼看就会成卖房笑话。 在中国没有一个行业的税、费象房地产那么名目繁多,方方面面都想分一杯羹。 为了提供公平竞争的市场环境,除特种行业外,其它各行各业所征税种、税率基本是一样的,即使有区别,包括特种行业,也至少对同一行业是一样的,唯独房地产是各地不同、五花八门,人为制造各种差异。中国房地产交易之复杂,十三亿人几乎没有一个第一次就能搞明白、一次过关的。这岂不违背最基本的市场原则?
http://ng1.17img.cn/bbsfiles/images/2016/11/201611141442_616419_0_3.jpg 近日,中国科学院上海应用物理研究所与清华大学合作,发展了一种新型的离子介导聚合酶链式反应(Ion-Mediated Polymerase Chain Reaction, IM-PCR),通过精确调控溶液pH值(即质子和氢氧根离子),可以在室温下完成PCR扩增,相关结果发表于《德国应用化学》(Angew. Chem. Int. Ed.)杂志,并申请了美国和国际专利。 PCR是体外通过酶反应合成、高灵敏扩增目标基因片段的一种方法,是最常用的分子生物学技术之一。常规PCR是通过控制溶液温度来实现DNA分子的可逆变复性,从而达到复制扩增的目的。而这种升降温过程通常需要复杂的控温装置并消耗大量能源。从理论上来说,DNA变复性还可以通过酸碱变性来实现,然而如何在不改变溶液体积的情况下精确控制溶液pH值则是一个难题。上海应物所物理生物学研究室樊春海课题组与清华大学化学系刘冬生课题组合作,建立了一种基于微流控的电化学芯片,可以通过电压驱动快速、精确调控溶液pH值,并实现了对DNA分子机器的电驱动控制(Nano Lett. 2010,10,1393)。基于这一研究基础,上海应物所博士张一和副研究员李茜发现微流控电化学可以有效控制PCR反应体系的pH值,实现了室温下的酸碱驱动的IM-PCR扩增。与传统PCR技术相比,这一新型的集成化IM-PCR技术无需变温过程,所有反应步骤均在室温下进行。这一小型化、低成本、易于集成的PCR新技术有望在生物检测、临床诊断及环境监测中发挥作用。
神经网络电活动增强快速调控抑制性突触稳态可塑性的分子机制 于翔研究组发表了题为“Postsynaptic spiking homeostatically induces cell-autonomous regulation of inhibitory inputs via retrograde signaling”的文章,文中阐述了神经网络电活动增强快速调控抑制性突触稳态可塑性的分子机制,这一研究成果公布在The Journal of Neuroscience杂志封面上。发育中的神经网络需要兼顾生长与稳定这两种相辅相成的需求。稳态可塑性可通过调节兴奋性或抑制性突触传递从而维持神经网络的稳定。已报道的关于稳态可塑性机制方面的研究主要集中在其对兴奋性突触传递的调节,很少关注其对抑制性突触的调控。研究人员发现,在体外培养的海马神经元中,持续增强神经元电活动4小时能够诱导抑制性突触传递的稳态上调,且这一过程明显早于兴奋性突触的变化。抑制性突触传递的稳态调节依赖于突触后神经元自身电活动的改变,是一种自我调节方式。这种调控通过突触后神经元分泌的脑源性神经营养因子(BDNF)逆突触作用于突触前的抑制性神经末梢,从而增强其自身的抑制性突触输入。重要的是,对幼年大鼠腹腔注射红藻氨酸,从而在体增强神经电活动,能够在海马CA1区域的锥体神经元中诱导出这种抑制性突触传递的稳态调控。这些结果提示,抑制性突触传递的自治性稳态调控是神经元应对网络电活动增强的一个快速代偿性保护反应。
来自美国Salk生物研究所,哥伦比亚大学等处的研究人员发表了题为“Inositol-1,4,5-trisphosphate receptor regulates hepatic gluconeogenesis in fasting and diabetes”的文章,发现了调控肝脏葡萄糖生成的分子开关,这也许为治疗胰岛素抵抗的II型糖尿病提供了一种新型治疗方法。相关成果公布在Nature杂志上。文章的第一作者之一是最新入选“青年千人计划”的王一国博士,王一国博士出国前在中科院上海生科院学习,后于Salk研究所做博士后,主要从事细胞质TORC调控机制和生理功能研究。荣柏生物I型糖尿病(旧称非胰岛素依赖型糖尿病或成人糖尿病)是一种代谢性疾病,其主要特征就是胰岛素抵抗,相对胰岛素缺乏和高血糖,这种疾病的患者体内产生胰岛素的能力并非完全丧失,有的患者体内胰岛素甚至产生过多,但胰岛素的作用效果却大打折扣。因此其中心问题确实就是葡萄糖生成的问题。在禁食状态,体液中循环的胰高血糖素的增加通过诱导糖原异生作用,促进了肝脏葡萄糖的产生。促发cAMP信号通路,通过CREB辅激活因子CRTC2的去磷酸化,提升了糖原异生基因的表达。胰高血糖素驴血清在一定程度上通过蛋白激酶A(PKA)介导的对CRTC2激酶SIK2的抑制作用,促进了CRTC2的去磷酸化。胰岛细胞能调控这种平衡,这些细胞位于胰腺中,能制造胰高血糖素,这是一种在禁食时候刺激肝脏产生葡萄糖,供给大脑使用的激素。这一过程在我们进食时会发生逆转,当胰岛细胞释放胰岛素的时候,这就能告诉肝脏停止制造葡萄糖。因此胰高血糖素与胰岛素是维持血糖稳定浓度的反馈组成部分。在这篇文文章中,研究人员识别出了一种延迟系统,解释了禁食阶段,葡萄糖如何激活CRTC2开关,以及这一系统在糖尿病中的作用。这一系统包含有一个肝细胞外侧分子受体:IP3——研究人员称之为“分子插口”,禁食阶段,胰高血糖素打开这一IP3插口,允许一个分子“油门”:一种钙调分子加速CRTC2的作用,激活基因,引导人血清 混合肝胆生成更多的葡萄糖,开启代谢引擎。研究人员还发现IP3受体的活性,以及肝脏中钙调磷酸酶 (calcineurin)在糖尿病胰岛素抵抗患者中会升高,导致更高血糖。因此可以通过选择性降低IP3插口,以及钙调磷酸酶这一加速器的活性,从而来关闭CRTC2开关,降低II型糖尿病患者的血糖浓度。
胆固醇稳态对机体正常的细胞和系统功能至关重要,胆固醇平衡失调是心血管疾病、神经退行性疾病和癌症等其他疾病的基础[1]。胆固醇代谢包括内源性胆固醇合成、吸收和排泄等环节。研究表明,胆固醇在体内不能被降解,有效排泄是维持其稳态的重要环节[2]。体内积累的胆固醇最终通过肠道以粪便消除胆固醇和胆汁酸的形式达到平衡,目前已知的胆固醇排泄途径包括了胆固醇逆转运(reverse cholesterol transport,RCT)途径和经肠胆固醇排泄(transintestinal cholesterol excretion,TICE)途径;前者是肝脏将胆固醇转化为胆汁酸后经肠排出,后者是由血直接经肠道分泌和排出血浆脂蛋白来源的胆固醇,二者交汇于肠道,因此,肠道在胆固醇稳态中发挥了重要作用[3-4]。调血脂治疗是防治体内高胆固醇含量诱导的相关疾病的有效方法,目前临床常用调血脂药物他汀类的作用是通过降低低密度脂蛋白胆固醇(low density lipoprotein cholesterol,LDL-C)以限制内源性胆固醇合成,从而防治心血管等疾病的发生发展,但其相关发病率和死亡率仅降低了30%[5]。这意味着需要更多策略来解决这个严重的公共卫生事件。 雷公藤红素(celastrol,CeT)是一种从传统中药雷公藤Tripterygium wilfordii Hook. f.中提取分离出的活性成分,具有抗炎、抗癌和抗动脉粥样硬化等多种药理活性[6],且具有良好的成药性,被《Cell》杂志列为最可能转化为现代药物的5种有潜力传统药物之一[7]。Zhang等[8]前期研究发现,体外有效成分为CeT的南蛇藤能够通过在促进RCT减少脂质蓄积方面具有积极作用,其机制主要是通过激活清除剂受体B类成员1(scavenger receptor class B member 1,SRB1)、ABC转运体和细胞色素P450家族7亚家族A成员1(cytochrome P450 family 7 subfamily A member 1,CYP7A1)途径促进胆固醇代谢。然而,有关CeT调控胆固醇代谢的作用机制探索尚不完善。此外,迄今为止,并无有关CeT通过调控肠道TICE途径介导胆固醇代谢的相关研究。因此,本研究采用网络药理学和系统生物学理论,通过构建“CeT-靶点-肠道胆固醇代谢”多层次网络,初步预测CeT调控肠上皮细胞胆固醇代谢的作用机制[9-10],并结合实验深入探讨和验证CeT调控肠上皮细胞胆固醇代谢的作用及机制,旨在为维持体内胆固醇稳态提供新的方向和理论依据。 1 材料 1.1 细胞 大鼠小肠隐窝上皮IEC-6细胞(批号ZQ0783)由中国科学院上海细胞库提供。 1.2 药品与试剂 CeT(批号C0869)购自美国Sigma公司;肝X受体α(liver X receptor α,LXRα)抑制剂GSK2033(批号HY-108688)、Bodipy荧光染色(批号HY-W090090)购自美国MCE公司;0.25%胰蛋白酶(批号PB180229)购自美国Hyclone公司;DMEM高糖完全培养基(批号ZQ-121)、DMEM基础培养基(批号09122)购自上海中乔新舟生物科技有限公司;磷酸酶抑制剂(批号CW2383S)、蛋白酶抑制剂(批号CW2200S)、BCA试剂盒(批号CW0014S)、SDS-PAGE试剂盒(批号CW0022S)、Loading buffer(批号CW0028S)液购自康为世纪生物科技股份有限公司;CCK-8试剂盒(批号C0037)、RIPA裂解液(批号P0013B)、ECL化学发光试剂盒(批号P0018S)购自碧云天生物技术股份有限公司;油红O染色试剂盒(批号G1262-4)购自北京索莱宝科技有限公司;PVDF膜(批号ISEQ00010)购自美国Millipore公司;兔抗三磷酸腺苷结合盒转运蛋白G5(adenosine triphosphate-binding cassette transporters G5,ABCG5)抗体(批号27722-1-AP)、甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体(批号10494-1-AP)、山羊抗兔二抗(批号SA00001-2)购自美国Proteintech公司;兔抗ATP结合盒转运蛋白G8(ATP-binding cassette transporters G8,ABCG8)抗体(批号A01482-2)购自武汉博士德生物工程有限公司;兔抗NPC1样细胞内胆固醇转运蛋白1(NPC1 like intracellular cholesterol transporter 1,NPC1L1)抗体(批号PA5-116672)购自美国Thermo Fisher Scientific公司;兔抗LXRα抗体(批号ab41902)、DAPI染液(批号ab228549)购自英国Abcam公司。 1.3 仪器 ix 73型倒置荧光显微镜(日本Olympus公司);FC型酶标仪、Forma 3系列CO2培养箱、EVOS fl auto全自动荧光倒置荧光学显微镜(美国Thermo Fisher Scientific公司);ChemiDoc XRS+化学发光成像系统、Mini-PROTEAN Tetra蛋白电泳系统(美国Bio-Rad公司)。 2 方法 2.1 网络药理学研究 将PubChem(https://pubchem.ncbi.nlm.nih.gov/)得到的CeT 3D结构导入PharmMapper(http://www. lilab-ecust.cn/pharmmapper/)进行药物靶点预测。通过NCBI(https://www.ncbi.nlm.nih.gov/)得到肠道和胆固醇代谢靶点。并与CeT靶点取交集得到共有靶点;通过STRING(https://STRING-db.org)进行蛋白质-蛋白质相互作用(protein-protein interaction,PPI)网络构建,并导入Cytoscape 3.9.1软件构建网络模型并分析;通过DAVID(https://david. ncifcrf.gov/)进行基因本体(gene ontology,GO)功能及京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析;利用PDB(https://www.rcsb.org/)筛选并下载分辨率小于0.25 nm的靶点的结晶复合式,结合上述得到的CeT 3D结构,采用Autodock进行分子对接,运用PyMol可视化处理。 2.2 实验验证 2.2.1 IEC-6细胞培养 IEC-6细胞用DMEM高糖完全培养基于37 ℃、5% CO2的恒温培养箱中常规培养。 2.2.2 CCK-8法检测细胞存活率 将对数生长期的IEC-6细胞接种于96孔板中,5×103个/孔。每孔加入100 μL含不同浓度(0.05、0.10、0.20、0.40、0.80 μmol/L)CeT的培养基,另设置加入无药物培养基的对照组,每组设置5个复孔,培养箱中培养24、48 h。每孔加入10 μL CCK-8试剂,于培养箱中孵育1~2 h后,采用酶标仪在450 nm处检测吸光度(A)值,计算细胞存活率。 细胞存活率=A给药/A对照 2.2.3 油红O染色评估CeT对肠上皮细胞胆固醇的影响 设置对照组、模型组和CeT(0.05、0.10、0.20 μmol/L)组,除对照组外,其余各组加入DMEM基本培养基配制的胆固醇胶束(cholesterol micelles, C-M,10 mmol/L)构建肠上皮细胞高胆固醇模型[11],给药组加入不同浓度的CeT溶液,对照组加入不含药物的培养基。干预24 h后,加入ORO Fixative固定液固定细胞;加入1 mL 60%异丙醇浸洗;加入油红O染液(ORO Stain A∶ORO Stain B=3∶2),洗涤至孔内无红色剩余;加入Mayer苏木素染色液,洗涤;加入油红O染液缓冲液;加入蒸馏水覆盖细胞并拍照,使用Image-Pro Plus软件以脂滴与整个图像的面积比进行定量。 2.2.4 Bodipy荧光标记法 按“2.2.3”项下方法进行分组和给药,干预24 h后,PBS洗涤,室温下多聚甲醛固定30 min;每孔加入2 μmol/L Bodipy染色液,于37 ℃细胞培养箱中孵育15 min;弃去染色液,PBS洗涤;DAPI复染核5 min,PBS洗涤;观察并拍照。使用Image-Pro Plus软件以荧光强度进行统计。 2.2.5 Western blotting检测TICE相关蛋白表达 按“2.2.3”项下方法进行分组和给药,干预24 h后,收集细胞;PBS洗涤后使用蛋白裂解液提取总蛋白质,BCA定量法测定蛋白质浓度。蛋白样品经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,转至PVDF膜,加入5%脱脂牛奶封闭3 h,用TBST洗涤后加入一抗,4 ℃孵育过夜;洗涤后加入二抗,室温孵育2 h;洗涤后进行显影,使用Image-Pro Plus软件分析条带灰度值。 2.2.6 免疫荧光检测LXRα/ABCG8和LXRα/ NPC1L1通路相关蛋白的影响 设置对照组、模型组、CeT(0.1 μmol/L)组、GSK2033(10 μmol/L)组和CeT+GSK2033组,除对照组外,其余各组加入DMEM基本培养基配制的胆固醇胶束(10 mmol/L)构建肠上皮细胞高胆固醇模型,各给药组加入相应药物,对照组加入不含药物的培养基。干预24 h后,PBS洗涤3次;4%多聚甲醛固定30 min,PBS洗涤3次后加入Trition X-100,室温下通透10 min;PBS洗涤3次后,加入免疫染色封闭液封闭60 min,吸去多余的牛血清白蛋白;分别滴加100 μL LXRα(1∶200)、ABCG8(1∶200)、NPC1L1(1∶200)一抗,4 ℃孵育过夜;回收一抗,PBS浸洗3次,滴加100 μL二抗(1∶300),室温避光孵育60 min;PBS洗涤3次,滴加DAPI复染核15 min,PBS洗涤3次;滴加抗淬灭剂10 μL,扣片,正面朝下盖在载玻片上,荧光显微镜下观察并拍照。使用Image-Pro Plus软件分析荧光强度。 2.2.7 统计学分析 采用GraphPad Prism 9.0和SPSS 26.0软件进行统计分析,数据以表示。两组间数据分布的正态性和方差齐性分别以Kolmogo? rov-Smirnov和Levene检验确定。组间均数比较采用t检验;多组间均数比较采用单因素方差分析,组间有差异进一步采用SNK-q检验进行两两比较。 3 结果 3.1 网络药理学研究 3.1.1 CeT-肠道胆固醇代谢靶点 通过TCMSP等数据库得到94个CeT相关靶点。通过NCBI Gene等数据库得到15 415个肠道相关靶点、14 177个胆固醇代谢相关靶点。并构建韦恩图预测CeT-肠道胆固醇代谢共有靶点,见图1。 图片 3.1.2 CeT-靶点-肠道胆固醇代谢网络构建 将PPI导入Cytoscape 3.8.1软件进行可视化,发现1个关键的子网络,见图2。 图片 3.1.3 CeT-肠道胆固醇代谢的GO功能富集分析 对CeT调控肠道胆固醇代谢的作用及机制进行GO富集分析,分别得到855个生物进程、17个细胞组成、53个分子功能,根据P<0.05,选出排名前10的条目,见图3。 图片 3.1.4 CeT-肠道胆固醇代谢的KEGG通路富集分析 CeT调控肠道胆固醇代谢的通路涉及34条,根据P<0.05,选出排名前10的通路,见图4。其中,主要涉及脂肪的消化和吸收、过氧化物酶体增殖物激活受体(peroxisome proliferators-activated receptor,PPAR)等信号通路。其中,肠道脂质代谢的关键通路(fat digestion and absorption)的靶点(ABCG5、ABCG8、NPC1L1)主要涉及CeT-靶点-肠道胆固醇代谢网络的关键网络之一(图5)。并且该网络主要涉及胆固醇排泄的关键途径——TICE途径。 图片 图片 3.2 分子对接分析 CeT与NR1H3(LXRα)结合能为?28.131 4 kJ/mol,可视化显示,匹配度良好,化合物与靶点结合的最优构象以氢键的方式呈现,结合活性良好,见图6。 图片 3.3 体外实验研究 3.3.1 CeT浓度筛选 不同浓度(0.05、0.1、0.2、0.4、0.8 μmol/L)的CeT分别干预IEC-6细胞24、48 h后,如图7所示,干预24 h时随着CeT浓度的增加细胞存活率降低,CeT的半数抑制浓度(half inhibitory concentration,IC50)为0.2 μmol/L。本研究在探索CeT安全浓度调控IEC-6细胞胆固醇代谢活性的同时,为了更进一步研究CeT在IC50时是否较安全浓度的效果更好,因此,选择0.05、0.10、0.20 μmol/L的CeT处理细胞24 h进行后续研究。 图片 3.3.2 CeT对IEC-6细胞内脂质的影响 如图8所示,油红O染色与Bodipy荧光标记结果均显示,与对照组比较,胆固醇胶束干预显著增加脂滴染色和荧光强度(P<0.001),表明造模成功;与模型组比较,各剂量CeT均显著抑制IEC-6细胞中脂质积累(P<0.05、0.01、0.001)。 图片 3.3.3 CeT对TICE途径关键蛋白的影响 NPC1L1是胆固醇吸收的重要蛋白,ABCG5/G8与胆固醇流出密切相关。如图9所示,与对照组比较,模型组NPC1L1蛋白表达水平显著升高(P<0.05),ABCG5和ABCG8蛋白表达水平显著降低(P<0.05、0.01);与模型组比较,CeT(0.2 μmol/L)组NPC1L1蛋白表达水平显著降低(P<0.05),CeT(0.1、0.2 μmol/L)组ABCG5和ABCG8蛋白表达水平显著升高(P<0.05、0.01)。因此,CeT可能通过抑制NPC1L1,促进ABCG5、ABCG8的表达,调控TICE途径介导的胆固醇摄取和流出。 图片 3.3.4 LXRα是CeT调控TICE途径的关键蛋白 如图10所示,与对照组比较,模型组LXRα、ABCG8表达显著降低(P<0.01),NPC1L1表达显著增加(P<0.001);与模型组比较,CeT组LXRα、ABCG8表达显著增加(P<0.001),NPC1L1表达显著降低(P<0.01),LXRα抑制剂GSK2033组ABCG8表达显著降低(P<0.01),NPC1L1表达显著增加(P<0.05);与CeT组比较,CeT+GSK2033组LXRα、ABCG8表达显著降低(P<0.01),NPC1L1表达显著增加(P<0.01)。因此,CeT可能通过促进LXRα的表达,调控TICE途径中的关键蛋白NPC1L1、ABCG8介导的胆固醇摄取和流出。 图片 4 讨论 胆固醇广泛存在于机体中,具有广泛的生理作用,是组织细胞中不可缺少的重要物质,它不仅参与细胞膜的形成,也是合成胆汁酸、维生素D及甾体激素的重要原料,但当其过量时便会导致高胆固醇血症,研究表明,心血管疾病、胆石症和肿瘤与高胆固醇血症密切相关[12-13]。胆固醇在体内不能被降解,机体有效排泄胆固醇是维持胆固醇稳态的重要环节[2]。因此,促进体内胆固醇排泄以维持体内胆固醇的动态平衡可能是治疗胆固醇失衡相关疾病的新策略。 基于此,本研究通过网络药理学方法,探讨CeT通过调控肠上皮细胞胆固醇代谢的潜在靶点及相关机制。PPI网络发现,CeT调控肠上皮细胞胆固醇代谢涉及1个核心子网络。其中,ABCG5/8与NPC1L1为胆固醇摄取与流出相关的核心靶点。本研究通过体外构建和模拟肠上皮细胞高胆固醇环境,探索CeT调控肠上皮细胞胆固醇代谢的机制,油红O和Bodipy结果均显示,CeT能够呈浓度相关性地降低胆固醇胶束干预的肠上皮细胞内的脂质积蓄。进一步通过结合KEGG通路分析发现,该子网络中的核心靶点与肠道脂质代谢的关键通路(fat digestion and absorption)相匹配,通过深度分析该通路发现,其主要涉及肠上皮细胞摄取与流出胆固醇中的TICE途径。已有研究表明,胆固醇从体内排出的唯一途径是通过粪便直接排出或转化为胆汁酸后排出,粪便排泄可通过2种独立途径进行,第1种途径是胆汁分泌,该途径已被广泛描述和研究。第2种途径是通过TICE途径[14]。在2009年Van团队初步研究估计,TICE对野生型小鼠体内排出的粪便中性固醇总量的贡献约为30%[15]。接下来,该团队在2010年通过实验得出在小鼠中TICE途径占粪便中性甾醇排泄的70%[16]。在人体生理情况下,TICE途径排泄的胆固醇占粪便胆固醇排泄总量的35%[2,17]。TICE指由血直接经肠道分泌和排出血浆脂蛋白来源的胆固醇。包括肝源性含载脂蛋白B的脂蛋白被基底膜侧低密度脂蛋白受体(low density lipoprotein receptor,LDLR)和其他可能受体吸收、内化,最终通过ABCG5/G8以及其他可能的转运体从顶端膜流出排泄到肠腔[18]。另有研究发现,利用Ezetimibe抑制NPC1L1介导的胆固醇摄取可显著增强TICE途径[2]。而本研究表明,CeT干预处于高胆固醇环境中的肠上皮细胞后,NPC1L1被抑制,而ABCG5、ABCG8被激活。提示,CeT主要通过抑制NPC1L1减少肠上皮细胞胆固醇摄取和促进ABCG5、ABCG8增加胆固醇流出。 LXRα由于其抗动脉粥样硬化、去除胆固醇和抗炎活性,在胆固醇稳态的转录调控中发挥极其关键作用[19]。研究发现,LXRα可调控NPC1L1在肠上皮细胞中的表达,降低肠道胆固醇的吸收[20]。此外,ABCG5和ABCG8是LXRα的直接靶基因,常形成异二聚体ABCG5/G8发挥作用,负责将细胞内胆固醇泵入肠腔并最终通过粪便排出体外[21]。PPI子网络表明,NPC1L1、ABCG5以及ABCG8主要由LXRα交联。因此,在上述研究的基础上,通过分子对接模拟了CeT与LXRα的对接模式,结合活性良好。采用LXRα抑制剂GSK2033处理,结果显示,NPC1L1和ABCG5/G8主要受LXRα调控。提示,CeT可能通过LXRα/ABCG5/ABCG8和LXRα/ NPC1L1途径分别介导IEC-6细胞胆固醇摄取和流出,进而促进TICE途径介导的胆固醇排泄。 本研究通过网络药理学和相关实验发现,CeT可能通过抑制肠上皮细胞胆固醇摄取和促进胆固醇流出维持机体胆固醇稳态,这一效应与核心靶点LXRα密切相关,本研究拓展了CeT调控体内胆固醇代谢的机制,为维持体内胆固醇稳态和胆固醇失衡相关疾病的新药研发提供了新思路。
菊苣酸是世界上广受欢迎的药用植物紫锥菊([b]Echinacea purpurea[/b] (L.) Menoch)的主要活性成分,被公认为商业热销紫锥菊产品的质量指标。虽然紫锥菊中菊苣酸的生物合成途径已被近期阐明,但其调控网络仍然不清楚。通过共表达和系统发育分析,该研究发现了[b]EpMYB2[/b],一个典型的R2R3型MYB转录因子(TF),对甲基茉莉酸甲酯(MeJA)刺激有响应,是菊苣酸生物合成的正调控因子。除了直接调控菊苣酸生物合成基因外,[b]EpMYB2[/b]还正向调控上游莽草酸途径的基因。我们还发现[b]EpMYC2[/b]可以通过结合其G-box位点激活[b]EpMYB2[/b]的表达,[b]EpMYC2-EpMYB2[/b]模块参与了MeJA诱导的菊苣酸生物合成。总的来说,我们鉴定了一个通过激活初级和特化代谢基因正向调控菊苣酸生物合成的MYB转录因子[b]EpMYB2[/b],连接了茉莉酸信号通路与菊苣酸生物合成之间的缺口。这项工作为利用生物技术手段提高紫锥菊药用质量开辟了新方向。 [align=left][b]前言[/b][/align] 几千年来,人类一直使用植物来维持健康。天然产物分子是药物开发的重要资源)。紫锥菊([b]Echinacea purpurea[/b] (L.) Menoch)原产于北美,并在全球广泛种植。除了作为观赏植物种植外,它还被用作草药治疗口疮、感冒和蛇咬伤。现代药理学实验已经证明它具有免疫调节、抗炎、抗氧化和抗病毒活性。紫锥菊产品几十年来在全球范围内畅销,通常用于预防和治疗普通感冒。在紫锥菊中发现了许多不同的化学成分,包括咖啡酸衍生物、糖蛋白、烷基胺和黄酮类化合物。其中,菊苣酸作为一种咖啡酸衍生物,是这些多种化学成分中最具代表性的,广泛积累在整个植物中,已被用作紫锥菊产品和原材料的质量指标。许多研究报告已经证明,菊苣酸具有多种生物活性,例如抗病毒、抗氧化、抗炎、肝脏保护、肾脏保护和抗癌,这些生物活性已经在最近的综述中得到了介绍。到目前为止,紫锥菊一直是菊苣酸补充剂的主要来源。由于市场对紫锥菊及其菊苣酸的需求量大,有必要提高植物中菊苣酸的含量,这依赖于生物合成途径和调控网络的阐明。 我们之前已经阐明了紫锥菊中菊苣酸的生物合成途径。首先,苯丙氨酸通过限速酶苯丙氨酸解氨酶(PAL)和随后的一些酶(如肉桂酸4-羟化酶(C4H)和4-香豆酸连接酶(4CL))转化为对香豆酰CoA。对香豆酰CoA是多种苯丙素类化合物生物合成的关键中间体,包括黄酮类化合物、羟基肉桂酸衍生物和白藜芦醇类化合物(Vogt, 2010)。在菊苣酸的生物合成过程中,对香豆酰CoA通过羟基肉桂酰CoA:莽草酸/奎宁酸羟基肉桂酰转移酶(HCT)和对香豆酰莽草酸/奎宁酸3'-羟化酶(C3'H)转化为咖啡酰CoA。两种BAHD型酰基转移酶(羟基肉桂酰CoA:酒石酸羟基肉桂酰转移酶(HTT)和羟基肉桂酰CoA:奎宁酸羟基肉桂酰转移酶(HQT))利用咖啡酰CoA作为酰基供体,分别以酒石酸和奎宁酸为酰基受体,生成槲皮素酸和绿原酸。最后,一种特殊的丝氨酸羧肽酶样(SCPL)型酰基转移酶——菊苣酸合酶(CAS)催化菊苣酸的生物合成,以槲皮素酸为酰基受体,绿原酸为酰基供体(Fu, Zhang, Jin等,2021)。我们进一步描述了酰基转移酶的底物多样性,并描绘了紫锥菊中菊苣酸及其类似物的整个生物合成网络。毫无疑问,菊苣酸的积累是由其生物合成基因的表达决定的。与此同时,生物合成基因的表达通常受到转录因子(TFs)的调控。尽管我们已经阐明了生物合成途径,但菊苣酸的调控网络仍然不清楚。 植物的各种转录因子(TFs)家族已经被发现参与植物次生代谢的调控,例如v-Myb髓样细胞白血病病毒癌基因同源物(MYB)、基础/螺旋-环-螺旋(bHLH)、WD重复、乙烯响应因子(ERF)、WRKY和基础亮氨酸拉链(bZIP)。其中,MYB转录因子被认为是苯丙素代谢的主要调控因子。它们包含一个保守的N端DNA结合结构域重复(R)和一个可变的C端调控区域。根据R的数量,MYBs可以分为四类,包括1R-、R2R3-、3R-和4R-MYB蛋白。R2R3-MYBs是植物MYBs中占主导地位的,被认为在功能多样化中帮助植物从水生环境适应到陆地环境。亚家族3、4、5、6、7、8、13、21、31、32、44和79的R2R3-MYBs主要被报道调控苯丙素生物合成途径,通过直接结合启动子作为激活因子或抑制因子发挥作用。许多植物R2R3-MYBs识别DNA靶点的AC元件,这些元件富含腺嘌呤和胞嘧啶残基。例如,AtMYB12,亚家族7的成员,被报道直接结合启动子的MYB12BS位点(CACCTACC、TACCTAMC和TAGCWACC),调控蔗糖磷酸合酶(DAHPS)、苯丙氨酸解氨酶(PAL)、查尔酮合酶(CHS)和黄烷酮3-羟化酶(F3H)的表达。 此外,生物和非生物刺激也影响植物次生代谢物的生物合成。植物防御机制的诱导通常伴随着大量次生代谢物的产生。在此过程中,脂质衍生植物激素——茉莉酸(JAs)发挥了关键作用。例如,JAs诱导了许多次生代谢物的生物合成,如长春碱、青蒿素、尼古丁和紫杉醇。许多转录因子家族的成员对JAs有响应,并调控JAs诱导的次生代谢物积累。在茉莉酸信号通路中,茉莉酸ZIM域(JAZ)家族的抑制蛋白通常与转录因子结合并抑制其激活功能。在JAs存在的情况下,JAZ蛋白被SCFCOI1复合物降解,最终释放转录因子。该通路中研究最透彻的转录因子属于MYC家族(bHLH型TF),包括MYC2、MYC3和MYC4。例如,MYC2s直接和间接地调控次生代谢物的诱导,通过结合下游基因启动子上的G-box位点并激活其表达。 有趣的是,菊苣酸的生物合成在紫锥菊的毛状根、幼苗和细胞悬浮培养物中显著受到甲基茉莉酸甲酯(MeJA)的诱导。目前尚不清楚哪种转录因子参与了紫锥菊中MeJA诱导的菊苣酸生物合成。将转录因子与生物合成基因结合使用将显著提高目标代谢物的产量。因此,必须识别潜在的正向调控因子以实现高产量的菊苣酸生产。最近,我们建立了紫锥菊开花期不同组织的转录组,并鉴定了一个属于亚家族6的R2R3-MYB转录因子[b]EpMYB1[/b],该转录因子正向调控紫锥菊中的花青素生物合成。在此基础上,我们通过共表达和系统发育分析鉴定了可能参与菊苣酸生物合成的转录因子。通过系统表征和体内转基因验证,鉴定出一个MYB转录因子,并阐明了其调控机制。进一步的研究还解释了其在MeJA诱导的菊苣酸生物合成中的作用。这项研究进一步阐明了菊苣酸的生物合成,并为利用生物技术手段提高紫锥菊中菊苣酸含量奠定了基础。 [align=left][b]结果[/b][/align] [b][b]参与菊苣酸生物合成的TF的筛选[/b][/b] 该团队之前已经阐明了紫锥菊中菊苣酸的生物合成途径(图1A)。利用最近建立的多组织转录组数据集,我们通过BLASTP方法识别出了菊苣酸生物合成基因。通过对这些生物合成基因表达水平的层次聚类分析,我们发现它们的表达模式相似,并且在根部高表达(图1B)。尤其是[b]EpPAL[/b]、[b]EpC4H[/b]、[b]Ep4CL[/b]、[b]EpC3’H[/b]、[b]EpHTT[/b]和[b]EpCAS[/b]具有紧密的共表达关系(图1B)。这些结果通过RT-q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]得到了验证。共表达分析已经被广泛应用于中间代谢物、生物合成基因和调控因子的鉴定。随后,我们使用这六个高度相关的菊苣酸生物合成基因作为诱饵进行共表达分析。通过与这六个基因共表达且线性相关系数高于0.8的基因的交集,共获得了139个转录因子(图1C)。MYB转录因子已经被证明可以调控苯丙素代谢。在这139个转录因子中,21个被注释为MYB。在去除冗余序列后,我们将这21个[b]EpMYBs[/b]与[b]AtMYBs[/b]进行比对并用于系统发育分析。这些[b]EpMYBs[/b]分布在几个亚家族中,包括2、7、14、20和78。之前有报道指出,亚家族3、4、5、6、7、8、13、21、31、32、44和79的成员参与调控苯丙素代谢。在这10个[b]EpMYBs[/b]中,亚家族7的两个成员(即CL4945.Contig5_All和CL4945.Contig7_All)可能具有调控苯丙素的潜力。当这两个[b]EpMYBs[/b]在[b]Nicotiana benthamiana[/b]叶片中瞬时过表达时,CL4945.Contig5_All(命名为[b]EpMYB2[/b])显著增加了总酚含量(图1D)。总体而言,我们通过共表达、系统发育分析和异源功能研究确定了[b]EpMYB2[/b]为潜在的菊苣酸生物合成调控因子。 [b][b]EpMYB2[/b] 正向调控菊苣酸生物合成[/b] 对[b]EpMYB2[/b]与拟南芥亚家族7成员的多序列分析表明,[b]EpMYB2[/b]具有完整的R2和R3结构域,可被鉴定为R2R3-MYB。进一步对[b]EpMYB2[/b]与其他植物物种亚家族7成员的系统发育分析显示,[b]EpMYB2[/b]与[b]Gentiana trifloral[/b]的[b]GtMYBP4[/b]较为接近,后者已被报道能促进类黄酮生物合成(图2A)。RT-q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]分析表明,[b]EpMYB2[/b]在根、茎和叶柄中高表达,这与RNA-seq数据一致(图2B)。通过在烟草叶片中瞬时表达[b]EpMYB2-GFP[/b]融合蛋白,发现[b]EpMYB2[/b]定位于细胞核中(图2C)。此外,[b]EpMYB2[/b]的表达对不同的胁迫处理(尤其是MeJA)具有响应(图2D),这表明它可能在环境胁迫与次生代谢之间起到链接作用。 为了探索[b]EpMYB2[/b]的体内功能,我们构建了[b]EpMYB2-OE[/b]紫锥菊愈伤组织。对照组和[b]EpMYB2-OE[/b]组愈伤组织的表型都呈现淡黄色,彼此之间没有显著差异(图3A)。[b]EpMYB2-OE[/b]组的[b]EpMYB2[/b]表达水平显著高于对照组(P 0.05)(图3B)。采用基于[url=https://insevent.instrument.com.cn/t/5p][color=#3333ff]液相色谱[/color][/url]-高分辨率质谱(LC-HRMS)的非靶向代谢组学方法,对紫锥菊愈伤组织进行代谢物变化评估。在主成分分析(PCA)得分图中,对照组与[b]EpMYB2-OE[/b]组显著分离(图3C)。进一步的载荷图评估表明,主要的差异离子属于菊苣酸及其类似物甲基菊苣酸和其底物槲皮素酸与绿原酸,根据保留时间和质谱进行鉴定(图3D)。与对照组相比,[b]EpMYB2-OE[/b]组中这些化学物质的含量显著增加(图3E)。此外,菊苣酸生物合成基因,包括[b]EpPAL[/b]、[b]EpC4H[/b]、[b]Ep4CL[/b]、[b]EpHCT[/b]、[b]EpC3’H[/b]、[b]EpHQT[/b]、[b]EpHTT[/b]和[b]EpCAS[/b],均受[b]EpMYB2[/b]上调(图3F)。这些结果表明,[b]EpMYB2[/b]是菊苣酸生物合成的正向调控因子。另一方面,鉴于亚家族7的成员被确认是类黄酮调控因子,我们进一步研究了[b]EpMYB2[/b]对类黄酮的影响。[b]EpMYB2[/b]对关键生物合成基因的表达和类黄酮代表性化学物质的水平没有表现出一致的促进作用。它显著增加了芸香苷的含量,但未显著增加紫茉莉苷的含量。菊苣酸的增加幅度远高于芸香苷,这表明[b]EpMYB2[/b]对菊苣酸生物合成的激活作用比对类黄酮的更强。所有这些结果表明,[b]EpMYB2[/b]主要在紫锥菊中促进菊苣酸的生物合成。 [b][b]EpMYB2[/b] 正向调控上游的初级代谢基因[/b] 许多转录因子被发现具有多个调控靶点,尤其是一些调控次生代谢的转录因子,它们也被发现参与初级代谢的调控。为了进一步评估[b]EpMYB2[/b]的功能,我们对转基因愈伤组织进行了RNA-seq分析。将上调的基因进行KEGG途径富集分析。令人感兴趣的是,除了苯丙氨酸代谢外,苯丙氨酸、酪氨酸和色氨酸的生物合成途径也得到了富集(图4A)。这些芳香族氨基酸来源于莽草酸途径,属于初级代谢途径,并且是苯丙素代谢的上游(图4B;表S1)。我们使用RT-q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url]评估了这些上游生物合成基因的表达水平。几种结构基因,包括[b]EpDAHPS[/b]、[b]EpEPSPS[/b]、[b]EpCM[/b]、[b]EpPAT[/b]和[b]EpADT[/b],被[b]EpMYB2[/b]显著上调(P 0.05)(图4C)。因此,色氨酸、酪氨酸和苯丙氨酸的含量均显著增加(P 0.05)(图4D)。综上所述,[b]EpMYB2[/b]被发现具有多个潜在的调控靶点,覆盖了初级和次级代谢途径。 [b][b]EpMYB2[/b] 通过直接结合激活关键的苯丙氨酸和菊苣酸生物合成基因的表达[/b] 为了探索[b]EpMYB2[/b]对菊苣酸生物合成的调控机制,我们尝试克隆这些初级和次级代谢基因的启动子。使用之前报道的方法,我们成功克隆了10个启动子(图5B)。然后,我们采用双荧光素酶测定法来测试[b]EpMYB2[/b]对这些启动子激活的作用(图5A)。[b]EpMYB2[/b]对[b]proEpCM[/b]、[b]proEpPAT[/b]、[b]proEpPAL[/b]、[b]proEpC4H[/b]、[b]proEp4CL[/b]、[b]proEpHCT[/b]、[b]proEpC3’H[/b]、[b]proEpHTT[/b]和[b]proEpCAS[/b]表现出激活效应,但对[b]proEpHQT[/b]没有影响(图5C)。在菊苣酸生物合成中起主导作用的关键代谢基因[b]EpPAL[/b]、[b]EpHCT[/b]和[b]EpHTT[/b]被广泛激活(图5C)。[b]EpHQT[/b]在紫锥菊种子发芽过程中的表达模式也表现出明显的差异,这表明其具有不同的调控网络。这些结果表明,[b]EpMYB2[/b]可以通过直接激活重要代谢基因的启动子来调控菊苣酸的生物合成。为了进一步识别潜在的结合位点,我们对这些启动子进行了分析,发现[b]proEpCM[/b]、[b]proEpPAT[/b]、[b]proEpPAL[/b]、[b]proEpHCT[/b]、[b]proEpC3’H[/b]和[b]proEpHTT[/b]在ATG上游500 bp内都包含[b]MYB12BS[/b]位点(图5B)。我们选择了[b]EpPAL[/b]、[b]EpHCT[/b]和[b]EpHTT[/b]这三个菊苣酸生物合成的关键代谢基因,来研究潜在的结合位点。在[b]EpPAL[/b]、[b]EpHCT[/b]和[b]EpHTT[/b]的启动子上突变[b]MYB12BS[/b]位点后,在双荧光素酶测定法中[b]EpMYB2[/b]对这些启动子的激活水平显著降低(图S7A;图5D、F)。此外,我们采用酵母单杂交(Y1H)实验来确认直接的结合效应。[b]EpMYB2[/b]显示出直接结合在[b]proEpHCT[/b]和[b]proEpHTT[/b]的[b]MYB12BS[/b]位点上(图5E、G)。由于[b]proEpPAL[/b]具有较高的自活性,因此难以验证[b]EpMYB2[/b]对其的直接结合作用(图S7B)。这些结果表明,[b]MYB12BS[/b]至少是[b]EpMYB2[/b]的一个结合位点。综上所述,我们发现[b]EpMYB2[/b]可以通过与启动子结合直接激活基因表达,并识别了一个结合位点。 [b][b]EpMYC2-EpMYB2[/b] 模块介导了MeJA诱导的菊苣酸生物合成[/b] 先前的研究表明,紫锥菊中菊苣酸的生物合成受到MeJA的诱导。在本研究中,我们注意到[b]EpMYB2[/b]的表达显著受到MeJA的影响(图2D),这让我们推测[b]EpMYB2[/b]是否参与了MeJA诱导的菊苣酸生物合成。由于[b]EpMYB2[/b]主要在根部表达(图2),我们处理了紫锥菊根部以进一步探索[b]EpMYB2[/b]在JA信号响应中的作用。结果表明,[b]EpMYB2[/b]的表达水平显著被MeJA处理所诱导(图6A)。此外,菊苣酸生物合成基因的表达水平和化学物质的含量也均受诱导(图6B)。考虑到[b]MYC2[/b]在JA信号途径中的重要作用,我们通过BLASTP分析发现了[b]EpMYC2[/b],该基因是从[b]Artemisia annua[/b]中鉴定出的[b]AaMYC2[/b]的同源基因。多序列比对分析表明,[b]EpMYC2[/b]包含完整的[b]MYC2[/b]结构域,包括基础和HLH(螺旋-环-螺旋)结构域、JAZ结合域(JID)以及与MED25蛋白结合的转录激活域(TAD)。系统发育分析显示,[b]EpMYC2[/b]与[b]AaMYC2[/b]较为接近,是bHLH亚家族7的典型成员(图6C)。为了探究[b]EpMYC2[/b]是否会影响[b]EpMYB2[/b]的表达,我们克隆了包含两个G-box位点的[b]EpMYB2[/b]启动子。[b]EpMYC2[/b]通过双荧光素酶测定法验证了其对[b]EpMYB2[/b]启动子的激活作用(图6D)。当启动子上距ATG 126 bp的G-box位点被突变后,[b]EpMYC2[/b]对[b]EpMYB2[/b]启动子的激活水平显著下降(图6D)。通过酵母单杂交(Y1H)实验确认了[b]EpMYC2[/b]直接结合[b]proEpMYB2[/b]–126 bp G-box位点的作用)。因此,[b]EpMYC2[/b]通过G-box位点激活了[b]EpMYB2[/b]启动子的表达。以上结果表明,[b]EpMYC2-EpMYB2[/b]模块介导了MeJA诱导的菊苣酸生物合成。 [align=left][b]结论[/b][/align]该研究发现了一种 MYB TF,即 EpMYB2,它能通过直接激活涵盖初级和特化代谢基因的初级和特化代谢基因的表达,正向调控紫锥菊中的菊苣酸生物合成通过直接激活涵盖初级和次级代谢的初级和特化代谢基因的表达,积极调控紫锥菊的菊苣酸生物合成。次生代谢基因的表达。鉴定EpMYC2-EpMYB2模块还连接了JA信号途径与菊苣酸生物合成之间的联系。这些结果进一步解释了菊苣酸的生物合成,并为菊苣酸的工程生产铺平了道路。此外,与 CK 组几乎不产生菊苣酸相比,EpMYB2 过度表达的紫锥菊中的菊苣酸含量相当可观,可将其视为提取菊苣酸的原料。
“中国中央政府网站”关于房地产市场调控的网上调查,让中央知道我们的疾苦http://www.gov.cn/diaocha/060602.htm大家顶~~~~~~,让中国的最高层知道我们的疾苦,让虚高的房价降下来。这非一般网站,而是中国中央政府网站,调查结果可能对中央的决策起一定的作用!!!
1.可以控制磨削量,一次磨削5-10微米。2.载荷可调控,最小载荷5牛顿左右。3.同时样品2-4个。4.在不从磨抛机上取下样品的状况下,可以通过监控设备(例如摄像头)观察样品表面磨抛状况。我们样品情况:大小:500-1000微米的陶瓷球材质:陶瓷球分若干层。每层的材质不同,包括SiC层和石磨层。电话:01-89796096 李
1.可以控制磨削量,一次磨削5-10微米。2.载荷可调控,最小载荷5牛顿左右。3.同时样品2-4个。4.在不从磨抛机上取下样品的状况下,可以通过监控设备(例如摄像头)观察样品表面磨抛状况。我们样品情况:大小:500-1000微米的陶瓷球材质:陶瓷球分若干层。每层的材质不同,包括SiC层和石磨层。联系电话:010-89796096 李老师
[font=仿宋_GB2312][size=21px]我国农田土壤的主要污染物有汞、镉、铅、铬、砷等重金属污染物,有机磷、有机氯等有机污染物,还有残留的农膜。[/size][/font][font=仿宋_GB2312][size=21px]农田土壤的污染途径包括农药、化肥和农膜的不正确使用;生活污水、商业污水、工业污水的不合理灌溉;矿业、工业固体废弃物在农田的不合理堆放;工业生产、汽车尾气排放及汽车轮胎磨损产生的大量含重金属的有害气体和粉尘在农田的自然沉降。[/size][/font]
[font=仿宋_GB2312][size=21px]我国农田土壤的主要污染物有汞、镉、铅、铬、砷等重金属污染物,有机磷、有机氯等有机污染物,还有残留的农膜。[/size][/font][font=仿宋_GB2312][size=21px]农田土壤的污染途径包括农药、化肥和农膜的不正确使用;生活污水、商业污水、工业污水的不合理灌溉;矿业、工业固体废弃物在农田的不合理堆放;工业生产、汽车尾气排放及汽车轮胎磨损产生的大量含重金属的有害气体和粉尘在农田的自然沉降。[/size][/font]
各市、县(区)生态环境局(分局)、财政局,银川市、石嘴山市、吴忠市、固原市、中卫市公共资源交易中心,宁东能源化工基地管委会生态环境局、财政金融局,国家税务总局宁夏区各市、县(区)税务局:为贯彻落实自治区党委十三届五次全会、自治区“六权”改革推进会精神和《关于深化“六权”改革的实施意见》(宁党办〔2023〕52号)有关要求,进一步规范和优化排污权储备和调控管理,现将《宁夏回族自治区排污权储备和调控管理办法》印发给你们,请遵照执行。《宁夏回族自治区储备和调控管理办法(试行)》(宁环规发〔2021〕5号)同时废止。[align=right]宁夏回族自治区生态环境厅 宁夏回族自治区财政厅[/align][align=right]宁夏回族自治区 国家税务总局[/align][align=right]公共资源交易管理局 宁夏回族自治区税务局[/align][align=right]2023年12月25日[/align](此件公开发布,联系人:纳静 5160939)[align=center][b]自治区排污权储备和调控管理办法[/b][/align]第一章 总 则第一条 为规范排污权储备和调控行为,完善排污权交易制度体系,根据《自治区关于开展排污权有偿使用和交易改革加快建设环境污染防治率先区的实施意见》《关于深化“六权”改革的实施意见》及其他相关规定,制定本办法。第二条 本办法适用于自治区范围内排污权储备和调控管理。法律法规规章另有规定的,从其规定。第三条 排污权储备是指自治区和设区的市政府(含宁东能源化工基地管委会,下同)通过无偿收回、有偿收储等形式,将排污权纳入政府储备的行为。生态环境等部门和单位(以下简称“排污权储备管理机构”)受政府指定,负责排污权储备和出让等具体工作。排污权出让是指排污权储备管理机构以协议出让、公开出让和小排放量项目简易出让等方式,将政府储备的排污权出让给排污单位的行为。排污权调控是指排污权储备管理机构根据排污权市场供需、环境质量状况和主要污染物总量控制目标任务,适时通过排污权交易系统回购或出让排污权,有针对性调节市场供求,保障重点项目建设的行为。第四条 自治区和设区的市建立财政保障机制,按照分级负责、属地管理原则,统筹资金用于回购排污权,支持排污权储备和调控。第五条 自治区和设区的市生态环境部门应会同财政、税务、公共资源交易等部门,建立排污权储备调控机制。生态环境部门负责排污权储备和出让的审查、核定及动态调控,编制排污权储备预算;同级财政部门根据年度预算规模拨付资金,用于排污权回购,并负责排污权储备和出让资金收支的监管;税务部门负责排污权出让收入的征收。第六条 自治区公共资源交易管理部门建设、运行和维护全区统一的排污权交易系统。第二章 排污权储备第七条 排污权储备的来源和方式:(一)通过市场交易或者协议出让等方式回购的排污权;(二)排污单位违反相关规定,被关停、取缔,不再排放相关污染物的,其无偿取得的排污权由排污权储备管理机构无偿收回,有偿取得的由排污权储备管理机构有偿收储;(三)新(改、扩)建项目自环评文件批准之日起5年内未开工建设或停止建设,其有偿获得的排污权指标确实无法实现排污权交易的,可由政府回购收储;其无偿获得的排污权指标(总量指标),由政府无偿收回;(四)新(改、扩)建项目在建设期间,因主动变更先进工艺技术或为适应污染物排放新标准进行提标改造,导致其有偿获得的排污权指标高于实际污染物排放量的部分,形成的富余污染物排放量,作为可交易排污权通过市场交易出售,确实无法实现交易的,可由政府回购收储;(五)排污单位可交易排污权自核定之日起5年内未使用或未交易的,可由政府收储;(六)由各级政府参与投资建设的集中式水污染治理设施、工业污染深度治理工程项目所减少的污染物排放量,扣除完成主要污染物总量减排任务减排量后剩余部分,按相应的投资比例纳入政府储备;(七)银行业金融机构处置排污单位抵押的排污权,可申请由政府回购;(八)其他符合法律法规规定的情形。第八条 排污权回购包括排污权储备管理机构主动回购和排污单位申请回购两种方式,具体程序如下:(一)排污权储备管理机构主动回购的,由排污权储备管理机构书面通知排污单位提供排污许可证、环评批复文件等相关材料;排污单位申请政府回购的,由排污单位向排污权储备管理机构提交排污权储备申请表,并按照来源类型提供排污许可证、环评批复文件等相关材料;(二)排污权储备管理机构收到申请材料后,对材料完整性、规范性进行审查。对符合储备范围、材料齐全的申请,予以受理;(三)排污权储备管理机构对受理的拟储备的排污权进行核定;(四)排污权储备管理机构按照有关审核意见向符合有偿储备条件的排污单位出具告知书;(五)排污权储备管理机构委托公共资源交易管理部门按照协议出让方式组织开展排污权回购交易,交易价格为排污权有偿使用费征收标准;(六)排污权储备管理机构向排污单位拨付交易资金,将回购排污权纳入储备;(七)生态环境部门为出让排污权的排污单位办理变更排污许可证。第九条 储备资格审核涉及现场检查的,根据管理权限进行,相关检查资料和检查结果应整理归档备查。第三章 排污权出让第十条 排污权储备管理机构应根据生态环境质量持续改善要求和排污权市场需要,对储备排污权实行动态调控。政府储备排污权自核定之日起有效期为五年,到期自动延续。第十一条 政府储备排污权的出让,原则上采用公开竞价方式进行交易。对战略性新兴产业、国家产业政策鼓励类和自治区、地级市(含宁东能源化工基地)优先培育发展的重大、重点扶持产业项目、民生保障类项目、污染物排放指标达到国家清洁生产标准一级水平的建设项目,及积极将可交易排污权投放市场的企业新(改、扩)建项目等,可按照协议出让方式进行交易,并给予优先保障和适当优惠。第十二条 来源于工业企业减排、破产、关停、淘汰、取缔的储备量,应优先投放市场。第十三条 工业、农业、服务业储备量原则上在本行业内交易;火电行业储备量优先在本行业内交易。第十四条 排污权储备管理机构应拟定排污权指标出让计划,并按照相关程序规则,以公开竞价或协议出让方式投放市场进行交易。公开竞价底价不得低于排污权基准价格;协议出让优惠额度由排污权储备管理机构根据实际情况确定。第十五条 排污权出让收入作为政府非税收入,纳入财政预算,收入列“1030715-排污权出让收入”,实行“收支两条线”管理,需要政府承担的必要支出纳入同级财政预算统筹安排。第十六条 政府储备排污权出让收入,自治区级部分全部缴入自治区级国库;地级市和宁东基地管委会储备排污权出让收入按照自治区与地级市、宁东基地管委会2:8比例分成。第四章 监督管理第十七条 排污权储备管理机构应真实、完整地记载和保存排污权储备调控的数量、价格和金额等有关资料,接受生态环境、财政、税务等部门的监督和检查。第十八条 相关单位工作人员有违规操作、弄虚作假、徇私舞弊、滥用职权行为的,依法依规严肃处理。第五章 附 则第十九条 排污权储备管理机构应定期向生态环境和财政部门报送辖区内排污权储备和调控及资金收支等情况。第二十条本办法由自治区生态环境厅、财政厅、公共资源交易管理局、国家税务总局宁夏回族自治区税务局负责解释。自2024年1月25日施行,有效期至2029年1月24日。宁夏回族自治区储备和调控管理办法(试行)》(宁环规发〔2021〕5号)同时废止。
加气机检定规程里有一句,计量调控装置必须加装封签或者电子封签,那么请问这个电子封签怎么定义?







 我要推广仪器
我要推广仪器
 下载APP
下载APP