[b]摘要[/b]从首次感染部位向邻近基质的转移入侵是肿瘤发展过程中的关键步骤,研究成果较少。肿瘤入侵的原理以各种体外模型给出了实验性的表述;但是,体内的关键性步骤和机制仍然不清楚。这里,我们通过落射荧光成像和多光子显微镜建立了一个修正的皮肤折叠室模型来阐述关于HT-1080纤维肉瘤细胞的原位移植,生长和入侵。这种策略允许对作为独立细胞或者集体粘丝或者细胞团沿着富含胶原的细胞外基质和增补宿主组织包括纹状肌肉丝和淋巴管的肿瘤生长、肿瘤诱导血管形成和入侵进行重复成像。这个修正的窗口模型将适用于阐述肿瘤转移和入侵的机制,以及相关的实验性治疗。[b]材料与方法[/b]HT-1080双色纤维肉瘤细胞表达细胞质DsRed2和核组蛋白2B(H2B)-EGFP -EGFP (Yamamoto et al. 2004)培养在改良的鹰培养基(PAN Biotech GmbH, Aidenbach, Germany)中,补充10%的胎牛血清(Aurion, Wageningen, The Netherlands),盘尼西林和链霉素(都100ug/ml PAN)和潮霉素B(0.2mg/ml;Invitrogen, Carlsbad, CA, USA)在37%湿润的5%CO2的培养环境中。小鼠被用异氟烷麻醉并被稳定固定在37℃的温控平台上。使用一个落射多光子显微镜[color=red]([/color][color=red]TriM Scope, LaVision BioTec[/color][color=red])[/color],并配备了OPO装置(OPO APE, Berlin, Germany)用于1100nm波段的双光子激发,以及红外修正的20X/0.95N.A(Olympus)物镜。如果没有特定声明,EGFP,DsRed2和SHG的获取都是使用的832nm的激发光。由带通滤波器确定的检测光波段为400/40(蓝),535/50(绿),605/70(红),和710/75(红外)。以5um的步长对深达250um的成像深度进行顺序3D堆栈。通过向尾静脉注射4mg荧光葡聚糖对血管显影。在注射了淋巴归巢环肽LyP-1(100ug)之后活化的淋巴管被检测到。(Laakkonen et al. 2002)图像被使用ImageJ 1.40 g (W. Rasband, NIH), ImSpector 3.4 (LaVision Bio- Tec GmbH), and Photoshop CS 8.0.1 (Adobe Systems Inc.)重构和分析。以宽的平方X长Xπ/6计算肿瘤体积。有丝分裂和细胞凋亡的比例通过H2B-EGFP模式从每区域30到100个细胞中确定。[b]主要结果 [/b][img=,593,498]http://qd-china.com/uploads/bio-product/51.jpg[/img]Fig.1 在背侧皮肤褶皱室中HT-1080纤维肉瘤细胞的滴落和注射方法比较.6(c)、7(d)天后通过明场和落射荧光显微镜观察的细胞应用,生长位置(a,b)和宏观肿瘤形态。在建立的模型中,允许细胞悬浮液或者细胞球粘附到外科手术准备好的真皮组织表面上,获得了在真皮层与盖玻片(a.c)间的3D肿瘤生长。使用细针将细胞球注射进真皮中阻止盖玻片和真皮内产量增加间的反应(b,d)。标尺1mm(概图)和250um(细节)。 [img=,604,379]http://qd-china.com/uploads/bio-product/52.jpg[/img]Fig 2. 肿瘤生长阶段。 a 由落射荧光显微镜监测的移植瘤生长和入侵的时间进程。新生血管的插入,不存在(3天)和存在(7天)。标尺1mm。b 通过以day 1的体积进行归一化的肿瘤体积。mean+-SD(n=9)。c HT-1080移植肿瘤在6天的时候的肿瘤形态,血管化,分生和凋亡。使用多光子显微镜以激发波长1100nm(左)和832nm(右)获取的一个中央中流区域的3D重构。核形态包括了有丝分裂(白色箭头)和凋亡图(黑色箭头)。标尺50um。插图显示了前相(P)、中相(M)和后相(LA)以及凋亡图(A)。d 对时间依赖的分生和凋亡定量化。数据显示3个非依赖性肿瘤的10-25个独立区域的Mean±SEM。 [img=,617,642]http://qd-china.com/uploads/bio-product/53.jpg[/img]Fig 3. 近红外多光子显微镜显示环绕HT-1080双色肿瘤的肿瘤诱导产生血管及其结构。Z轴为一个6天大肿瘤的从肿瘤边缘(-50um)到肿瘤内部区域(-80um)(红色细胞质;黄色细胞核)。通过FITC-葡聚糖注射现实的密布血管(绿),先前存在的线形血管(绿色箭头)和不规则形状的新生血管(蓝色箭头)。胶原纤维(黑色箭头)和肌肉丝(白色箭头),通过二次谐波检测(灰度)。标尺50um。 [img=,583,768]http://qd-china.com/uploads/bio-product/54.jpg[/img]Fig 4. HT-1080双色细胞的原位入侵模型。a 注射后6天入侵类型的分类。缺少入侵(上,左)并且散布单个细胞(上,右;白色箭头),散射的或者紧密地丝状整体入侵(下图)。标尺250um。 b 45个连续的非依赖性肿瘤的按中所分入侵模式的频率。11天时,沿着纹状肌肉纤维集体入侵丝的定位。标尺100um。d 单一细胞侵入脂肪组织随后进行分散的,部分整体的入侵。对照-少量圆的脂肪细胞(星号)被HT-1080细胞包围。1100nm的激发光来检测遍布的血管(Alexa Fluor 660-dextran,红色),,肿瘤细胞质(绿色假彩),SHG(灰度);832nm用于肿瘤细胞核(白色)。标尺100um。[img]http://qd-china.com/uploads/bio-product/55.jpg[/img]Fig 5. HT-1080细胞沿淋巴管的入侵。a 由多光子显微镜对边缘而非肿瘤中心的活化淋巴管产生的单幅图片。用FITC连接的LyP-1缩氨酸来检测。深度已标明在图上(um)。b 3D堆栈投影表明淋巴管内(白色箭头)和外淋巴管入侵(黑色箭头)。标尺100um。
[size=14px] [/size] [size=14px]青蒿素(Arteminsinin)是从植物青蒿中分离出来的倍半萜内酯,与它的一些衍生物一起被公认为一种有效的用于治疗疟疾药物,现已逐渐被认为是潜在的抗肿瘤药物,已有一些研究试图确定青蒿素的蛋白质靶点并破译青蒿素杀死癌细胞的分子机制,但迄今为止,青蒿素的确切抗肿瘤相关靶点仍有很大挖掘空间。[/size] [size=14px] [/size] [size=14px]1、细胞毒性筛查将ART1确定为潜在的抗肿瘤药物[/size] [size=14px]作者首先制备了C-10位的不同芳基取代基的青蒿素衍生物(ART1、ART2和ART3),利用肺癌细胞系H1299和A549比较了它们以及青蒿素(QHS)及其衍生物双氢青蒿素(DHA)的抗肿瘤活性。发现ART1,一种含有萘环的青蒿素衍生物,对肺癌细胞表现出最强的细胞毒性。在肿瘤类器官模型和白血病MV4细胞中均证明ART1是最有效化合物。此外,ART1表现出对正常细胞的抗增殖活性非常弱。结果表明ART1是一种有前途的潜在抗癌药物。[/size] [size=14px]图片[/size] [size=14px]图1 ART1抑制肿瘤生长[/size] [size=14px]2、ART1诱导非经典铁死亡[/size] [size=14px]先前的报告表明青蒿素通过多种方式导致癌细胞死亡,包括细胞凋亡、自噬等。作者发现ART1触发的细胞死亡与凋亡、自噬无关。进一步确定ART1诱导癌细胞死亡的机制,发现ART1诱导的细胞死亡仅被铁死亡抑制剂ferrostatin-1(可防止脂质过氧化物的积累)抑制,而不能被细胞凋亡抑制剂z-VAD-FMK或坏死性凋亡抑制剂necrostatin-1抑制,表明ART1处理触发铁死亡。此外,ART1处理会诱导脂质过氧化,且ART1引起的脂质过氧化是铁依赖性的。深入机制研究发现ART1导致铁死亡已知类别的铁死亡诱导剂不同,它不影响其细胞内GSH水平和GPX4活性。[/size] [size=14px]图片[/size] [size=14px]图2 ART1诱导非经典铁死亡[/size] [size=14px]3、鉴定HSD17B4蛋白作为ART1的直接靶标[/size] [size=14px]为了确定ART1介导诱导铁死亡的蛋白靶点,作者设计了并合成了ART16(生物素标记的ART1)来开展Pulldown。ART16类似于ART1可诱导铁死亡,可用于后续实验。Pulldown+蛋白质组学分析显示HSD17B4蛋白为可能靶点, BLI、Pulldown+WB技术证实了两者的直接结合。[/size] [size=14px]图片[/size] [size=14px]图3 鉴定HSD17B4蛋白作为ART1的直接靶标[/size] [size=14px]4、ART1通过HSD17B4蛋白介导癌细胞死亡[/size] [size=14px]作者采用ART99(含有香豆素荧光团的ART1探针),发现ART99与靶蛋白HSD17B4的共定位。通过敲低HSD17B4来研究ART1诱导的细胞死亡是否由HSD17B4介导,发现HSD17B4敲低可显著减弱ART1的作用。[/size] [size=14px]图片[/size] [size=14px]图4 ART1通过HSD17B4蛋白介导癌细胞死亡[/size] [size=14px]5、ART1靶HSD17B4蛋白直接诱导脂质氧化[/size] [size=14px]HSD17B4蛋白是一种双功能酶,同时具有脱氢酶和水合酶活性,并参与VLCFA(极长链脂肪酸)的过氧化物酶体β氧化。作者发现ART1并未改变细胞中HSD17B4蛋白丰度,也不影响其脱氢酶和水合酶活性。由于ART1中的过氧化物部分对于诱导铁死亡是必不可少的,作者推测ART1可能是一种启动铁死亡的选择性氧化剂,与HSD17B4结合并促进周围脂质的氧化。作者验证发现ART1可以直接氧化铁死亡相关底物。PUFA,易受脂质过氧化的影响,是执行铁死亡所必需的。由于不容易获得超长链多不饱和脂肪酸,AA被用作替代物,作者发现ART1单独可以氧化AA,ART1还可以显著促进由亚铁离子催化的脂质过氧化。此外,活细胞成像探针发现ART1可以氧化细胞中HSD17B4蛋白周围的脂质。这些数据证实ART直接氧化HSD17B4蛋白周边的脂质,积累脂质过氧化物,并最终在癌细胞中促进铁死亡。[/size] [size=14px]图片[/size] [size=14px]图5 ART1靶向HSD17B4蛋白直接诱导脂质氧化[/size] [size=14px]6、ART1优先诱导高间充质状态癌细胞的铁死亡[/size] [size=14px]据报道,高间充质状态的耐药性癌细胞对铁死亡诱导剂敏感。作者检测这些肺癌细胞的上皮间充质状态,发现对ART1敏感细胞系H1299和H1838中的波形蛋白含量较高,表明高间充质状态,而对ART1耐药细胞系HCC366和H1650几乎表现出E-钙粘蛋白的丰度检测不到,这表明ART1的敏感性与癌细胞上皮间充质状态密切相关,ART1可优先诱导间充质癌细胞发生铁死亡。[/size] [size=14px]图片[/size] [size=14px]图6 ART1优先诱导高间充质状态癌细胞的铁死亡[/size] [size=14px]总结[/size] [size=14px]该研究将青蒿素衍生ART1已被确定为铁死亡诱导剂,对癌细胞增殖具有显著的抑制效果。接着使用化学蛋白质组学方法鉴定HSD17B4蛋白,一种在VLCF分解代谢中必不可少的酶,作为ART1的直接靶点。进一步研究发现ART1会导致铁死亡,通过直接氧化HSD17B4蛋白周围的脂肪酸而不干扰蛋白质的正常酶活性,揭示了一种意想不到的机制,其中ART1-HSD17B4用作“特洛伊木马”,潜入过氧化物酶体触发脂质氧化。总之,ART1通过靶向HSD17B4诱导铁死亡提供了一种有希望的癌症治疗方法。[/size]
[b][size=15px][color=#595959]2型神经纤维瘤病(NF2)[/color][/size][/b][size=15px][color=#595959]是一种罕见的遗传综合征,易使个体发展为双侧[b]前庭神经鞘瘤(VSs)[/b],可导致高风险的危及生命的神经并发症。NF2相关VS的传统治疗方案通常会导致神经损伤,迄今为止,[/color][/size][b][size=15px][color=#595959]没有[/color][/size][size=15px][color=#595959]FDA[/color][/size][size=15px][color=#595959]批准的NF2药物疗法[/color][/size][/b][size=15px][color=#595959]。该研究的目的是评估中药复方[b]祛毒散结汤(QDSJ)[/b]对NF2相关的VS的[b]抗肿瘤疗效[/b],并研究潜在的[b]作用机制[/b]。[/color][/size] [align=center] [/align] [size=15px][color=#595959]超高效[url=https://insevent.instrument.com.cn/t/5p][color=#3333ff]液相[/color][/url][b]色谱-质谱[/b](UHP[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url])分析用于鉴定QDSJ的成分及其靶点。为了确定QDSJ的假定靶点与NF2相关VS的差异基因之间的关系,使用UHPL-MS数据结合前期的基因表达谱数据筛选[b]药物-疾病交叉基因[/b]。将差异表达的基因导入字符串数据库以生成PPI网络。使用GO和KEGG途径富集分析鉴定[b]差异表达的基因靶点和途径[/b]。[/color][/size] [size=15px][color=#595959]分别使用[b]患者来源的神经鞘瘤细胞系和患者来源的异种移植物小鼠模型[/b],检测QDSJ的疗效。H&E染色、[b]免疫[/b]组化和免疫荧光染色用于评估细胞增殖和肿瘤[b]血管[/b]。[/color][/size][font=mp-quote, -apple-system-font, BlinkMacSystemFont, &][size=15px][color=#595959][/color][/size][/font][align=center] [/align] [size=15px][color=#595959]通过UHP[url=https://insevent.instrument.com.cn/t/Yp][color=#3333ff]LC-MS[/color][/url]分析,共鉴定出133个化合物。网络药理学表明,[b]坏死、凋亡、细胞周期、血管生成、粘附连接和神经活性配体受体相互作用[/b]的调节可能与QDSJ治疗NF2相关VS的疗效相关。[/color][/size] [b][size=15px][color=#595959]QDSJ治疗可诱导坏死细胞死亡和神经鞘瘤细胞凋亡[/color][/size][/b][size=15px][color=#595959],并抑制肿瘤生长。组织病理学分析显示QDSJ处理的肿瘤中有细胞坏死区域和肿瘤血管扩大。与对照肿瘤相比,QDSJ治疗的肿瘤中细胞周期蛋白D1和Ki-67阳性的细胞数量显著减少。CD31和αSMA的免疫荧光染色显示QDSJ治疗的肿瘤中肿瘤血管数量和密度减少,血管结构正常化。[/color][/size][font=mp-quote, -apple-system-font, BlinkMacSystemFont, &][size=15px][color=#595959][/color][/size][/font][font=mp-quote, -apple-system-font, BlinkMacSystemFont, &][size=15px][color=#595959][/color][/size][/font][size=15px][color=#595959][/color][/size][color=#3573b9]结论[/color][b][size=15px][color=#595959][/color][/size][/b][font=mp-quote, -apple-system-font, BlinkMacSystemFont, &][size=15px][color=#595959][/color][/size][/font] [b][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][/b][size=15px][color=#595959]该研究表明,QDSJ对NF2相关神经鞘瘤具有[b]显著的抗肿瘤活性[/b],是未来临床试验的可能[b]候选药物[/b]。[/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size][size=15px][color=#595959][/color][/size]
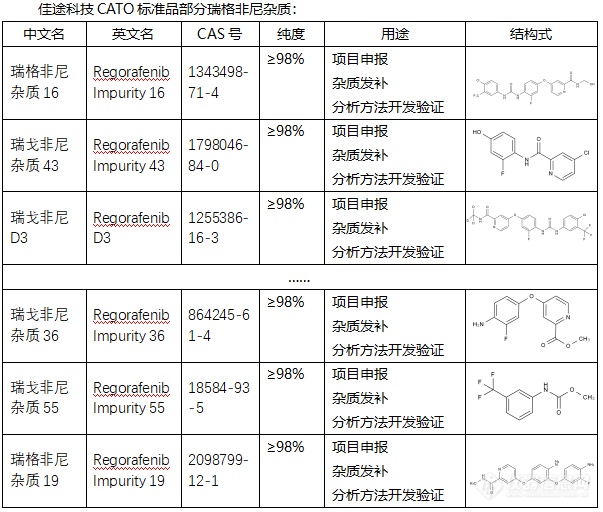
[font=宋体]◇关于瑞格非尼杂质[/font][font=宋体][color=#1f1f1f]瑞格非尼杂质[/color][/font][font=宋体][color=#1f1f1f]是[/color][/font][font=微软雅黑]一种多激酶抑制[/font][b][font=微软雅黑]杂质,它的作用机制主要是[/font][/b][font=微软雅黑]通过抑制多种蛋白质激酶[/font][font=微软雅黑]和[/font][font=微软雅黑][color=#666666][back=#f2f2f2]干扰肿瘤细胞的生长和进化来发挥作用[/back][/color][/font][font=微软雅黑][color=#666666][back=#f2f2f2],[/back][/color][/font][font=微软雅黑][color=#666666][back=#f2f2f2]从而[/back][/color][/font][font=微软雅黑][color=#333333]减少了生长因子对肿瘤细胞的[/color][/font][font=微软雅黑][color=#333333]作用[/color][/font][font=微软雅黑][color=#333333],[/color][/font][font=微软雅黑][color=#333333]达到[/color][/font][font=微软雅黑][color=#333333]抑制了细胞的生长和分裂[/color][/font][font=微软雅黑][color=#333333]的作用。它主要有以下四个作用:一[/color][/font][font=Arial][color=#333333][font=宋体]、[/font][/color][/font][font=宋体][color=#333333]抑制酪氨酸激酶[/color][/font][font=宋体][color=#333333];[/color][/font][font=宋体][color=#333333]二阻断肿瘤生长信号传导,抑制了肿瘤细胞的增长;三[/color][/font][font=Arial][color=#333333][font=宋体]、[/font][/color][/font][font=宋体][color=#333333]诱导肿瘤细胞凋亡;四[/color][/font][font=Arial][color=#333333][font=宋体]、[/font][/color][/font][font=宋体][color=#333333]抑制肿瘤的血管的生成,降低肿瘤细胞对血液供应的依赖。[/color][/font][font=宋体][color=#333333] [/color][/font][font=宋体][font=Calibri]CATO[/font][font=宋体]标准品提供的瑞格非尼杂质[/font][/font][font=微软雅黑],在治疗一些类型的癌症上具有十分显著的疗效。[/font][img=,603,512]https://ng1.17img.cn/bbsfiles/images/2024/02/202402042208499344_2683_6381607_3.png!w603x512.jpg[/img]
请问谁有新出的标准《塑料一差示扫描量热法(DSC)第六部分:氧化诱导时间和氧化诱导温度的测定》?能不能分享一下?谢谢!
1.目的了解外源基因在原核细胞中表达的特点和方法。2.原理外源基因克隆在含有lac启动子的表达系统中。先让宿主菌生长,lac I产生的阻遏蛋白与lac操纵基因结合抑制下游的外源基因转录。向培养基中加入诱导物IPTG(异丙基硫代-b-D-半乳糖),解除抑制使外源基因大量表达。表达的蛋白可经SDS-PAGE或Western-blotting检测。3.器材旋涡混合器,微量移液取样器,移液器吸头,50ml 微量离心管,1.5ml 微量离心管,双面微量离心管架,台式冷冻离心机,制冰机,恒温摇床,分光光度计,超净工作台,恒温培养箱,摇菌试管,三角烧瓶,接种环。4.试剂LB培养基(加抗菌素),100mg/ml IPTG,20%葡萄糖。5.实验准备无菌ddH2O,1.5ml离心管装入铝制饭盒(灭菌)、移液器吸头装入相应的吸头盒(灭菌),牙签(灭菌),摇菌管(灭菌),100mg/ml IPTG (过滤灭菌)(100ml分装,-20°C保存),100mg/ml氨苄青霉素(过滤灭菌)(100ml分装,-20°C保存),配制20%葡萄糖(8磅灭菌20分钟,添加至上述LB中,终浓度为0.2%)。6.操作步骤(1) 晚上9:00接种。在超净工作台中接种含有Pinpoint™xa-3-CHI重组载体的菌株,培养于两个三角烧瓶各20ml LB-葡萄糖培养基(含抗菌素Amp 120μg/ml)的摇菌管中,慢速70~90转/分钟30°C摇菌过夜。(2) 至第二天上午8:30 OD600约为0.5,加IPTG至 100μg/ml,150~170转/分钟37°C诱导1.5-3h。同时做不加IPTG诱导和非转化的空菌诱导的对照培养。(3) 4000rpm离心15min弃掉上清液,收获菌体,用SDS-PAGE电泳分析(表达蛋白分子量为30kDa)。菌体也可放在-20°C以下保存备用。(4) 在被细菌污染的桌面上喷洒70%乙醇,擦干桌面。
哺乳动物雷帕霉素靶蛋白(mTOR)是一种丝/苏氨酸蛋白激酶,于1991年被Heitman等研究啤酒酵母细胞突变体抵抗雷帕霉素毒性作用中发现并提出[1]。雷帕霉素是一种由大量蛋白质组成的大环内酯类药物,属于磷酸肌醇3-激酶相关蛋白激酶(PIKK)家族。mTOR与癌症有密切关系[2-3],mTOR信号转导通常参与调节细胞的存活、生长、代谢、蛋白质合成和自噬、稳态[4]。mTOR有两种不同的多蛋白复合物mTORC1、mTORC2。mTORC1对雷帕霉素敏感,激活参与mRNA翻译的p70S6激酶(p70S6K)和真核起始因子4E结合蛋白(4E-BP1)。mTORC2被认为对雷帕霉素有耐药性,通常对营养和能量信号不敏感[5]。mTORC1受磷脂酰肌醇3激酶/蛋白激酶B(PI3K/Akt)通路、Ras/Raf/MEK/ERK通路和其他细胞内因子等多种信号通路调节[4, 6]。目前对mTORC2的研究较少,有研究表明mTORC2通过磷酸化AGC激酶,包括Akt、蛋白激酶C(PKC)和血清/糖皮质激素调节激酶1(SGK-1)来发挥作用[7]。mTOR上游信号传导通路主要由PI3K/Akt等介导。mTOR下游信号通路为p70S6K、4E-BP1等,通过促进其翻译和蛋白质合成的磷酸化来介导[8]。目前许多mTOR抑制剂被开发用于癌症的治疗[9]。一些中药活性成分可以通过mTOR信号通路促进细胞凋亡和自噬性死亡、抑制细胞增殖,发挥抗肿瘤作用[10-11]。目前关于中药活性成分抑制mTOR信号通路的研究较多,根据结构不同可分为蒽醌类、生物碱类、萜类、多糖类、黄酮类、多酚类成分。本文总结了中药活性成分抑制mTOR通路抗肿瘤作用的研究进展,明确其作用机制,为临床应用提供参考。 1 蒽醌类成分 蒽醌类化合物是一类具有良好抗癌作用的三环类天然有机化合物,其中2个酮基位于中心环,这种三环双酮核心结构具有特定靶向作用。如通过不同的上游途径靶向自噬,包括Akt/mTOR轴等从而达到抑癌结果[12]。Zhang等[13]研究发现大黄酸诱导口腔癌细胞中的活性氧(ROS)积聚以抑制Akt/ mTOR信号传导通路,通过Akt/mTOR信号通路诱导口腔癌细胞凋亡和ROS在体内外发挥抗癌作用。另有研究发现大黄酸联合mTOR抑制剂依维莫司作用胃癌细胞MGC-803,可抑制p-磷脂酰肌醇3激酶(PI3K)、p-Akt和p-mTOR的表达发挥协同抗肿瘤作用[14]。此外,大黄酚与mTOR抑制剂雷帕霉素组合时,通过表皮生长因子受体(EGFR)/mTOR介导的信号转导途径显著阻断细胞增殖[15]。研究发现紫九牛总蒽醌通过下调p-Akt、p-mTOR蛋白的表达而抑制PI3K/Akt/mTOR信号通路活化抑制胃癌细胞SGC7901细胞的增殖、诱导凋亡、抑制迁移和侵袭的能力[16]。 2 生物碱类成分 生物碱含有环状结构,其中至少1个碱性氮原子被并入其中,广泛分布于豆科、防己科、毛茛科等植物中[17]。生物碱及其衍生物种类众多,具有相似环结构,经不同代谢途径合成的生物碱可能具有不同的药理活性。郝艳梅等[18]观察到苦参碱培养的人非小细胞肺癌A549细胞造成细胞萎缩、碎裂显著增加,可以观察到自噬液泡,在加入PI3K特异性抑制剂后发现可以减少p-Akt和p-mTOR的表达,诱导A549细胞自噬和凋亡增加,说明苦参碱通过抑制PI3K/Akt/mTOR通路实现。Zhang等[19]发现小檗碱可以通过PI3K/Akt通路逆转小鼠黑色素瘤B16细胞的上皮间质转化,起到参与治疗黑色素瘤的作用。另有研究发现,小檗碱通过PI3K/Akt/ mTOR途径调节人甲状腺未分化癌细胞的自噬和凋亡[20]。Li等[21]发现小檗碱抑制Notch 1通路导致PTEN表达增加,进而下调PI3K/Akt/mTOR通路,导致直肠癌细胞SW480细胞周期停滞和自噬发生,产生抑制肿瘤细胞增殖的作用。另外的研究中发现小檗碱可通过诱导结直肠癌细胞Ht-29、Sw-480和Hct-116凋亡和坏死来抑制细胞的增殖、迁移和侵袭,通过上调PTEN、下调PI3K、Akt和p-Akt的表达和抑制其下游靶点mTOR、p-mTOR来调节PI3K/Akt通路的活性[22]。苦参碱和小檗碱已经发挥出了抑制肺癌、甲状腺癌等的抑制作用,更多的生物碱类成分在mTOR信号通路的作用亟待被发现。 3 萜类成分 萜类化合物由异戊二烯或异戊烷单元以各种方式连接在一起,并具有不同类型的闭环、不饱和度和官能团,可分为单萜、倍半萜、二萜、三萜等,多样的结构为抗肿瘤药物的开发提供了较多的选择性[23]。Jang等[24]研究发现从泽泻分离的三萜类成分表现出抗肿瘤活性,如泽泻醇A、泽泻醇B、泽泻醇B 23-乙酸酯等。在1项研究中,乳腺癌MDA-MB-231细胞p-Akt、p-mTOR和p70 S6K的表达水平在泽泻醇A处理后显著下调,表明泽泻醇A可以抑制PI3K/Akt/mTOR信号传导[25]。同样在乳腺癌细胞MCF-7、雌激素受体阳性乳腺癌MCF-7细胞中,穿心莲内酯通过下调雌激素受体α(ERα)、PI3K和mTOR的表达水平抑制细胞增殖[26]。除以上萜类成分外,其他如白桦脂醇对转移性结直肠癌细胞的抗增殖作用[27]、柴胡皂苷A联合化疗药物促进前列腺癌细胞的死亡和缓解耐药[28]、银杏内酯抑制肝细胞癌[29]、土贝母皂苷甲诱导乳腺癌细胞自噬激活Akt活性化合物[30]、土贝母总皂苷抑制三阴性乳腺癌细胞MDA-MB-231细胞的增殖[31]均与各种mTOR相关信号通路的调控机制有关。以上研究表明中药活性成分单独应用或联合应用时可以作为治疗癌症的潜在Akt/mTOR抑制剂。 4 多糖类成分 中药多糖具有抗肿瘤、抗病毒、抗氧化等活性,可有效抑制肿瘤细胞增殖分化[32]。多糖的抗肿瘤活性与其一级结构、高级结构有关,其中每种因素对多糖的抗肿瘤活性都有不同程度的影响。Yao等[33]从枸杞多糖进一步提取和分离具有短肽骨架和复杂的分支聚糖部分的肽聚糖(LbGP),研究发现其能抑制癌细胞生长,还可以通过蛋白激酶A-cAMP反应元件结合蛋白(PKA-CREB)通路促进PER2的表达,而PER2抑制PI3K/Akt/mTOR通路负性调节固醇调节元件结合蛋白-1c(SREBP1c)的表达抑制胶质母细胞瘤中的脂质合成,从而抑制胶质母细胞瘤细胞增殖。研究者发现黄芪多糖抑制结直肠癌细胞HCT-116细胞和小鼠肿瘤组织中PI3K/Akt/ mTOR信号通路相关蛋白p-PI3K/PI3K、p-Akt/Akt、p-mTOR/mTOR的表达诱导自噬,从而减少肿瘤细胞的生长[34]。通过对宫颈癌U14荷瘤小鼠的实验研究提示半枝莲多糖可能通过抑制mTOR/p70S6K信号通路使凋亡基因Bcl-2表达减少,促进细胞凋亡,发挥抑瘤作用[35]。另有研究发现山慈菇多糖抑制肝癌腹水荷瘤小鼠肿瘤生长[36],黄芪多糖导致肿瘤细胞细胞程序性死亡-配体1(PD-L1)表达降低以增强化疗效果[37],这些也是通过mTOR信号通路调控自噬、免疫等达到的治疗目的。可见枸杞多糖、黄芪多糖、山慈菇多糖等中药多糖均可通过调控mTOR通路对肝癌、结直肠癌等肿瘤细胞有着体内、体外的抑制作用。 5 黄酮类成分 黄酮类化合物的基本母体是由2个具有酚羟基的苯环通过3个碳原子相互连接而成的C6-C3-C6单元[38]。抗肿瘤活性程度与各类黄酮母核结构差异、C-2,3位是否存在双键等的化学结构有密切关系[39]。异槲皮苷是存在于杨梅等植物中的黄酮类化合物,Shui等[40]发现使用异槲皮苷处理的人肝癌细胞HepG 2、Huh 7通过激活AMPK/mTOR/p70S6K途径触发自噬诱导细胞死亡,而异槲皮苷触发失调的自噬促进caspase依赖的凋亡性细胞死亡。小豆蔻素是从草豆蔻中分离得到的查耳酮,Jin等[41]研究发现小豆蔻素可以抑制三阴性乳腺癌细胞株MDA-MB-231的生长,是通过抑制mTOR/p70S6K通路从而在mRNA和蛋白水平抑制低氧诱导因子-1α(HIF-1α)的表达,进而增强线粒体氧化磷酸化,诱导ROS的积累达到抑癌作用的。更多研究如荔枝核总黄酮通过抑制Akt/mTOR等信号通路诱导前列腺癌细胞(PCa)凋亡,抑制PCa细胞的体内生长和体外增殖、转移[42]。异甘草素在体内和体外通过诱导自噬有效地抑制肝癌细胞的增殖,并诱导凋亡,可能通过PI3K/Akt/mTOR通路[43]。桑根醇L在前列腺癌细胞中抑制PI3K/Akt/mTOR信号传导诱导其凋亡[44]。羟基红花黄色素A通过抑制肝癌细胞PI3K/Akt/mTOR通路触发自噬反应抑制肿瘤细胞生长[45]。研究中发现异鼠李素可降低MAPK14的表达,抑制胃癌细胞HGC-27细胞的增殖和迁移,促进其细胞凋亡,进一步研究显示异鼠李素通过调节MAPK/mTOR信号通路抑制胃癌细胞增殖[46]。另有研究提示黄酮类化合物对肺癌、乳腺癌、结直肠癌等的抗癌作用中有着积极的表现[47]。以上结果说明黄酮类化合物可对抗恶性肿瘤的发生、发展,对应用于临床、解决实际问题有一定的潜力。 6 多酚类成分 多酚类是一类由1个或多个直接连接到芳族烃基的有机化合物。外界因素的诱导使多酚在原有的结构上经羟基化、甲氧基化、脱糖基化、单体聚合等结构修饰,从而发挥各种药效作用。经结构修饰后的多酚类成分往往具有更高生物活性、更好临床疗效[48-49]。在1项实验中发现芦荟素的使用抑制了肝细胞癌HepG2、Bel-7402细胞的增殖和侵袭,进一步研究发现其通过激活PI3K/Akt/mTOR途径诱导肝细胞癌的凋亡和自噬[50]。在另一项研究中发现芦荟素可抑制胃癌细胞HGC-27、BGC-823 GC细胞增殖和迁移,可能是通过抑制烟酰胺腺嘌呤二核苷酸磷酸氧化酶2(NOX2)的激活来抑制ROS的产生,从而抑制Akt/mTOR等信号通路的磷酸化[51]。Zhang等[52]发现毛兰素可能通过抑制PI3K/ Akt/mTOR通路实现诱导肺癌细胞凋亡、G2/M期阻滞,抑制其迁移和侵袭,在体内实验中减少肿瘤组织的血管比率、增加凋亡肿瘤细胞的数量、上调白细胞介素(IL)-2和肿瘤坏死因子-α(TNF-α)水平等对肿瘤细胞的积极抑制作用。此外,红景天苷通过PI3K/Akt/mTOR通路抑制胃癌生长,并诱导细胞凋亡和保护性自噬[53]。姜黄素通过抑制Akt/mTOR通路抑制肾癌细胞ACHN细胞活力,诱导凋亡和自噬[54],通过修饰关键基因和蛋白的表达下调PI3K/ Akt/mTOR信号通路抑制头颈肿瘤细胞的增殖[55],以及对雄激素依赖性和非依赖性前列腺癌的抑制作用[56]。白藜芦醇调节NGFR/AMPK/mTOR信号通路诱导肺癌细胞A549细胞自噬和凋亡[57]。仙鹤草素显著破坏线粒体功能,降低mTOR/HIF-1α通路蛋白表达,影响细胞内能量代谢,诱导胰腺癌细胞凋亡[58]。其他酚类如石斛酚[59]、6-姜烯酚[60]均可调节mTOR信号通路抑制癌细胞的增长。学者对于酚类化合物的研究较多,这为其尽早应用于临床提供了实验依据。 7 结语 mTOR是重要的信号传导通路,在癌细胞中被过度激活,使肿瘤细胞增殖,抑制该通路可以起到一定抗肿瘤作用。蒽醌类、生物碱类、萜类等中药活性成分可以通过抑制mTOR通路诱导肿瘤细胞凋亡、促进自噬、阻断细胞周期,逆转上皮间质转化等一系列抑癌作用对肺癌、胃癌、结直肠癌等肿瘤细胞起到治疗作用。中药活性成分抑制mTOR信号通路的国内外研究展现了其在改善患者生存质量,降低耐药、减少患者复发率等方面良好的前景。中药活性成分具有不良反应较小、抗肿瘤作用明显的优点,但也有不足之处,如中药靶向性差、生物利用度不足、消除速度快等。目前靶向mTOR信号通路作为先导化合物研发出高效、安全的中药抗肿瘤新药的研究还很欠缺,对此应该进行更深入的研究。不过随着相关领域的基础理论、实验和技术手段的不断更新,相信不久会将精准靶向mTOR信号通路的中药活性成分选出,并明确其作用机制,将基础实验结果推向临床应用。
肿瘤是指体内细胞的异常增生,可以是良性的或恶性的。良性肿瘤(例如息肉)生长缓慢且通常局限在一个区域,不会侵犯周围组织或扩散到其他部位。恶性肿瘤(即癌症)具有侵袭性,可以快速生长并通过血液或淋巴系统扩散到其他身体部位,形成远处转移。癌症是一种严重威胁人类健康和生命的疾病,2020年全球有1 930万新增癌症病例和1 000万癌症死亡病例,且我国癌症发病率和死亡率均位居全球第一[1]。最常见的癌症类型是乳腺癌、肺癌、结直肠癌和前列腺癌。因此,寻找新的抗肿瘤药物,阐明抗肿瘤药物的分子机制,是解决当前临床肿瘤治疗难点的有效策略。中药具有多种有效成分,因其不良反应低、多靶点、多通路等优点,已成为抗肿瘤药物开发的重要来源和研究热点[2]。目前,常规的肿瘤症治疗方法为手术、放射治疗和化学治疗等,但这些方法往往伴随着较大的不良反应和毒性,而且对某些难治性或复发性肿瘤效果不佳[3]。因此,寻找有效、低毒的抗肿瘤药物是当前临床研究的重要方向。 苍术是一种常用的中药材,分为茅苍术Atractylodes lancea (Thunb.) DC.和北苍术A. chinensis (DC.) Koidz.,分别来源于菊科植物茅苍术或北苍术的干燥根茎。苍术具有燥湿健脾、祛风散寒的功效,在《神农本草经》中列为上品[4]。近年来,苍术在抗微生物、抗炎、抗肿瘤、免疫调节、调节消化系统、心血管系统和神经系统等方面的药理作用受到了广泛关注。苍术中含有挥发油、多糖、倍半萜类、聚乙炔类等[5]多种化学成分。其中一些成分已经被证实具有抑制或杀伤多种肿瘤细胞的能力,其作用机制涉及诱导凋亡、抑制增殖、迁移、侵袭和转移,以及调控免疫功能等方面[6]。然而,苍术中的抗肿瘤活性成分及其作用机制尚未完全明确,需要进一步深入地探索和验证。本文通过整理国内外研究文献,对苍术活性成分、苍术与其他药物联合抗肿瘤及其分子机制进行总结,探讨苍术在抗肿瘤方面的应用规律和思路,为苍术资源的开发利用以及抗肿瘤临床疗法的研究提供理论参考。 1 苍术主要化学成分 茅苍术与北苍术化学成分相似,药理作用也较为相似,目前已从苍术中分离出多种化学成分,主要含有包括萜类、聚乙烯炔类、有机酸类、糖苷类化合物等[7-8]。苍术主要抗肿瘤化学成分,见图1。茅苍术与北苍术中主要化学成分如表1所示。 图片 图片 2 苍术的抗肿瘤机制 苍术中含有苍术内酯Ⅰ、Ⅱ、Ⅲ、苍术酮、β-桉叶醇和苍术素等有效成分,这些成分不仅可以抗炎、抗氧化、抗菌、保肝、降血糖,还可以抗肿瘤[14-15]。近年来,苍术及其有效成分对肿瘤的抑制作用受到了广泛的关注。研究发现,苍术有效成分对多种肿瘤细胞都有抑制作用,可以通过多种途径和机制影响肿瘤细胞的生长、迁移、侵袭和血管生成,诱导肿瘤细胞的凋亡和自噬,调节肿瘤微环境和免疫系统。 2.1 抑制肿瘤细胞增殖 肿瘤是由于细胞增殖失控而形成的一种疾病[16]。细胞周期是细胞增殖的基本过程,由细胞周期蛋白(cyclin,CCN)和细胞周期蛋白依赖性激酶(cyclin-dependent kinase,CDK)复合物共同调控[17]。干预细胞周期是抑制肿瘤发展的有效策略之一[18]。Kotawong等[19]发现,苍术中的苍术素、苍术内酯I和β-桉叶醇等有效成分可以通过影响肿瘤细胞周期的不同阶段来抑制肿瘤细胞的增殖。这些成分可以通过抑制磷脂酰肌醇-3-羟激酶(phosphatidylinositol 3-hydroxy kinase,PI3K)、磷酸化蛋白激酶B(protein kinase B,AKT)和哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路来诱导肿瘤细胞在G1期停滞;Yu等[20]发现苍术内酯I通过上调周期蛋白依赖性激酶抑制剂1A(cyclin-dependent kinase inhibitor 1A,p21)和下调cyclinB1、CDK1和细胞分裂周期25C蛋白(cell division cyclin25,Cdc25c)等关键分子来抑制肿瘤细胞在G2/M期的进入,在动物模型中,苍术内酯I可以显著抑制膀胱癌的生长,且无明显不良反应。Zhang等[21]实验发现苍术内酯Ⅱ可以通过改变结直肠癌细胞内的蛋白表达从而抑制结直肠癌细胞的增殖和活性,并且还显著增强了结直肠癌细胞的化疗敏感性。Pongsakorn等[22]发现,苍术提取物可以通过抑制细胞外信号调节激酶信号级联(ERK-signaling cascade,ERK)信号通路来抑制胆管癌细胞的增殖。ERK信号通路是一种重要的细胞内信号转导机制,参与调节细胞生长、分化和凋亡等过程。苍术提取物可以下调ERK及其下游分子的表达,从而抑制胆管癌细胞的生长和增殖,不同类型的胆管癌细胞对苍术提取物的敏感度不同,其中人胆管HuCCT-1癌细胞最为敏感。 2.2 诱导肿瘤细胞凋亡 细胞凋亡是一种程序性细胞死亡形式,它通过限制细胞的增殖和分化来维持组织稳态或去除潜在的有害细胞[23]。目前已知的细胞凋亡途径主要有3种,即外源性途径(死亡受体介导)、内源性途径(线粒体介导)和内质网途径。其中,线粒体途径是最重要的一种,它涉及线粒体外膜透化(outer mitochondrial membrane,MOMP)、细胞色素C释放和半胱天冬酶(cysteine aspartic acid protease,Caspase)激活[24]。多项研究发现,苍术酮可以通过降低线粒体膜电位、提高活性氧水平、抑制B细胞淋巴瘤-2基因(B-cell lymphoma-2,Bcl-2)表达、促进BCL2-相关X蛋白(BCL2-associated X protein,Bax)裂解和Caspase-3表达[25],以及下调PI3K/AKT/mTOR信号通路来诱导肿瘤细胞凋亡[26]。Narahara等[27]研究表明,β-桉叶醇和苍术内酯Ⅲ[27]可以通过增加Caspase-3、Caspase-8、Caspase-9和Bax等凋亡相关蛋白的表达、下调Bcl-2表达、释放细胞色素C和降低线粒体膜电位来诱导胆管癌细胞凋亡。此外,Li等[28]使用β-桉叶醇处理的白血病HL60细胞,发现β-桉叶醇可以通过激活c-JunN端激酶(c-Jun N-terminal kinase,JNK)丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号通路来诱导白血病HL60细胞凋亡。Li等[29]研究发现,苍术素可以通过降低Bcl-2表达、激活p53肿瘤蛋白(p53 tumor protein,p53)、Bax和Caspase-3、-8、-9等凋亡因子来诱导人乳腺癌MCF-7细胞凋亡,并表现出浓度依赖的毒性效应。Li等[30]研究表明,苍术内酯I和苍术内酯Ⅱ[31]可以通过与对两面针激酶2(Janus kinase 2,JAK2)直接相互作用而负调节信号传导及转录激活因子3(signal transducer and activator of transcription 3,STAT3)磷酸化,从而抑制其活化,进而导致糖酵解的抑制和结肠、直肠癌细胞凋亡的诱导。 2.3 抑制肿瘤细胞转移 肿瘤细胞转移是指肿瘤细胞通过血液循环从原发部位转移到其他部位的过程,这是癌症治疗的难点,也是癌症死亡的主要原因[32]。上皮间质转化(epithelial-mesenchymal transition,EMT)是一种与癌症发生相关的细胞程序,它使癌细胞具有移动性、侵袭性和抗凋亡能力,从而促进转移。苍术的一些活性成分具有抑制肿瘤细胞转移的潜在作用,其机制可能涉及对EMT的调控[33]。Acharya等[34]研究发现,β-桉叶醇可以改变EMT相关标志物的表达,从而抑制结肠癌细胞的增殖、迁移和侵袭。同时它还可以影响PI3K、AKT、p38丝氨酸/苏氨酸蛋白激酶(p38 mitogen-activated protein kinase,p38MAPK)信号通路,以及肺癌细胞中的活性氧水平,从而降低癌细胞的黏附和迁移能力[35]。麦静愔等[36]发现苍术酮可以通过抑制EMT过程等途径抑制肿瘤细胞的迁移和侵袭能力,此外,苍术酮还可以通过下调基质金属蛋白酶(matrix metalloproteinase,MMP)的表达从而抑制肿瘤细胞的迁移和侵袭能力。MMP是一类能够降解细胞外基质(extracellular matrix,ECM)的锌依赖性内肽酶,在癌症进展中的作用与它们参与ECM降解以及黏附和细胞骨架蛋白、生长因子、趋化因子的调节和加工有关[37]。且有动物实验表明,苍术酮可以明显抑制肝癌生长,没有明显的毒性。Zhong等[38]在观察了苍术多糖在U-2 OS人骨肉瘤细胞中对内皮细胞选择素(endothelial cell selectin,E-Selectin)和路易斯X三糖(Lewis-X Trisaccharide,LacCer Lex)的影响,发现苍术多糖可通过降低U-2 OS细胞上的E-Selectin抑制U-2 OS细胞对人脐静脉内皮细胞HUVECs的黏附、迁移和侵袭。肿瘤相关巨噬细胞(tumor-associated macrophages,TAMs)在促进肿瘤转移中发挥重要作用,Zhang等[39]发现苍术内酯II可以有效抑制肿瘤细胞极化,从而抑制肺癌细胞在体内和体外的转移。铁死亡是一种新的细胞死亡模式,其特征是铁过载导致脂质过氧化而导致膜损伤,过度的铁死亡会影响肿瘤的转移,从而抑制肿瘤的进展[40]。He等[41]发现,苍术素可通过抑制谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)和铁蛋白轻链(ferritin light chain,FTL)的表达,以及上调酰基辅酶A合成酶长链家族成员4(acyl-CoA synthetase long-chain family member 4,ACSL4)和转铁蛋白受体(transferrin receptor,TFR1)的表达来诱导肝癌HCCM细胞的铁死亡。 2.4 诱导肿瘤细胞自噬 细胞自噬是一种分解代谢通路,能清除不必要的或功能失调的细胞成分并回收代谢底物[42]。目前已知有3种主要的细胞死亡方式:细胞凋亡(Ⅰ型)、自噬性细胞死亡(Ⅱ型)和坏死(Ⅲ型)。自噬性细胞死亡是指自噬过程中产生的自噬体过多或过大,导致细胞质溶解和细胞死亡。自噬体是由双层膜包裹的囊泡,内含被降解的细胞器和蛋白质。微管相关蛋白1轻链3(microtubule-associated protein 1 light chain 3,LC3)是自噬体形成的关键标志物,它以微管相关蛋白1A/1B-轻链3(microtubule-associated protein 1 light chain 3,LC3-I)和微管相关蛋白1轻链3的脂化形式(lipidated form of microtubule-associated protein 1 light chain 3,LC3-Ⅱ)2种形式存在,LC3-Ⅰ转化为LC3-Ⅱ是自噬体形成的必要步骤[43-44]。Li等[29]使用苍术素处理乳腺癌MCF-7细胞时发现,苍术素可以增加了LC3Ⅰ向其脂化形式的LC3Ⅱ的转化,并增加了苄氯素1(beclin-1,BECN1)的表达,下调了人乳腺癌MCF-7细胞中的p62蛋白(p62 protein,p62)表达,改变凋亡和自噬相关生物标志物。Acharya等[45]研究发现,苍术素通过调节PI3K、AKT、mTOR、p38MAPK信号通路的活性,可以诱导胆管癌HuCCT-1细胞发生自噬,并抑制其生长、迁移和侵袭,SB202190(p38MAPK诱导剂)和3-MA(p38MAPK抑制剂)分别显著增加和降低苍术素诱导的自噬速率。 2.5 抑制肿瘤血管生成 血管生成本身不会导致恶性肿瘤的形成,但可以为肿瘤的生长和转移提供条件。肿瘤在发展到一定阶段后,需要依赖新生血管来满足其对氧气和营养的增加的需求,以及排除代谢废物,因此,抑制血管生成是一种有效的抗肿瘤策略[46]。血红素加氧酶1(heme oxygenase 1,HO-1)是一种在肿瘤组织中高表达的酶,它可以促进肿瘤的血管生成和抗氧化应激,为肿瘤细胞提供生存优势。因此,抑制HO-1的表达或活性是治疗肿瘤的另一种有效策略之一。Mathema等[47]研究发现,苍术素可以抑制胆管癌CL6肿瘤细胞的集落形成和伤口愈合能力,其机制与抑制HO-1的表达、下调信号转导及转录激活蛋白1/3(signal transducer and activator of transcription 1/3,STAT1/3)和核因子κB(nuclear factor kappa-B,NF-κB)的信号通路有关。β-桉叶醇也具有抑制胆管癌细胞中HO-1的表达的能力,其机制与浓度依赖性地抑制STAT1/3和NF-κB信号通路有关[48]。β-桉叶醇还可以通过抑制生长因子信号通路中的环磷腺苷效应元件结合蛋白(cyclic-AMP response binding protein,CREB)激活来阻断血管生成,从而抑制肿瘤的发展[49]。Tsuneki等[50]有动物实验表明,β-桉叶醇可以通过激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)来刺激大鼠嗜铬细胞瘤细胞中的神经突生长,且β-桉叶醇还表现出了体外和体内的抗血管生成活性,其阻断了由碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)或血管内皮生长因子(vascular endothelial growth factor,VEGF)诱导的人脐静脉内皮细胞(human umbilical vein endothelial cell,HUVEC)中CREB蛋白的磷酸化,从而抑制bFGF刺激的HUVEC迁移和HUVEC在基质胶中的管形成。同时,它还能显著降低小鼠皮下植入的Matrigel栓塞和小鼠佐剂诱导的肉芽肿中的血管生成[51]。 2.6 免疫调节作用 随着肿瘤的发生和发展,或在接受化疗、放疗等治疗的过程中,肿瘤患者机体免疫力的显著下降。因此,调节或刺激机体免疫能力,可能是一种有效的主动抗癌策略。免疫治疗作为一种新型的抗癌手段,已经引起了广泛的关注和研究[52]。巨噬细胞是机体内重要的免疫细胞,在机体免疫中发挥着重要的作用[53]。Qin等[54]从苍术中分离得到两种多糖成分:中性多糖和酸性多糖。研究表明,酸性多糖能够显著地刺激小鼠单核巨噬细胞白血病细胞(RAW264.7)细胞的增殖、吞噬能力、NO产生和细胞因子分泌,并且呈现出剂量相关性,而中性多糖则相对较弱。此外,中性多糖和酸性多糖均能够激活淋巴结Peyers patch细胞中的T细胞,并促进集落刺激因子的产生。而酸性多糖也表现出比中性多糖更好的肠道免疫调节活性。吲哚胺-2,3-二氧化酶(indoleamine 2,3-dioxygenase,IDO)是一种通过犬嘌呤途径氧化分解色氨酸的限速酶,是抗肿瘤免疫治疗中小分子药物开发的潜在目标。IDO可在肿瘤微环境中通过与许多肿瘤相关的自发炎症和T细胞激活而被诱导。Liu等[55]研究发现,苍术内酯Ⅰ可以通过下调Toll样受体4/髓样分化蛋白2复合物(toll-like receptor 4/myeloid differentiation 2 complex,TLR4/MD-2)的表达,抑制人卵巢癌细胞(EOCSKOV3)中髓样分化主要反应蛋白88(myeloid differentiation primary response protein 88,MyD88)、NF-κB、Akt和IDO1的信号通路的活化,从而减少白细胞介素-6(interleukin-6,IL-6)、转化生长因子-β1(transforming growth factor beta 1,TGF-β1)、VEGF和白细胞介素-17A(interleukin-17A,IL-17A)等促进肿瘤免疫逃逸的因子的分泌。同时,还可以降低调节性T细胞(Treg细胞)在肿瘤微环境中的比例,改善T淋巴细胞受到EOCSKOV3细胞上清液抑制而导致的增殖反应降低和抗肿瘤细胞毒性减弱。Liu等[56]研究发现,苍术内酯Ⅲ可以通过直接结合JAK3蛋白,从而抑制γ-干扰素(interferon gamma,IFN-γ)触发的JAK3/STAT3通路,从而达到抑制IDO激活的目的。 苍术抗肿瘤成分的潜在分子机制见图2。对苍术抗肿瘤有效成分及其抗肿瘤作用进行归纳总结,见表2。 图片 图片 3 联合用药 西医治疗肿瘤的常用手段有手术切除、药物化疗和高能射线放疗等,这些手段去除肿瘤西医的治疗方式更为直接,适合前期控制病情,化疗药物虽然能够杀死肿瘤细胞,但同时也伴有严重的副作用,影响患者的生活质量和治疗效果。中药具有不良反应小、安全性高的特点,因此中药与化疗药物的联合应用被广泛关注和探索[57]。 阿帕替尼是全球第一个在晚期胃癌被证实安全有效的小分子抗血管生成靶向药物,也是晚期胃癌标准化疗失败后,明显延长生存期的单药。Zhou等[58] 分析了不同苍术多糖提取方法的影响。比较了热水浸提法、超声浸提法和酶浸提法提取苍术多糖的得率、总糖含量、相对分子质量分布、单糖组成、并测定苍术多糖与阿帕替尼的协同活性。结果发现其中超声浸提法表现出最强的协同作用。这也与超声浸提的苍术多糖相对分子质量小、β-构型高、半乳糖含量高的事实相一致。Srijiwangsa等[59]发现,β-桉叶醇可以通过抑制胆管癌细胞和细胞裂解物中的NAD(P)H醌氧化还原酶1[NAD(P)H quinonedehydrogenase 1,NQO1]的活性和蛋白表达,增强氟尿嘧啶和多柔比星对细胞迁移的细胞毒性活性和抑制活性。Mai等[60]将不同浓度的苍术内酯I、硼替佐米以及硼替佐米+苍术内酯I作用于U266细胞结果研究发现,苍术内酯可以调节JAK2/STAT3通路上的IL-6、JAK2、STAT3等基因表达抑制U266肿瘤细胞的增殖和促进其凋亡并呈剂量依赖性,并能与硼替佐米产生协同作用,当苍术内酯I与硼替佐米联合使用时,可显著增强对U266细胞增殖的抑制作用。 紫杉醇是第一个获得批准的草药衍生化疗药物[61]。并且作为一种已知的Toll受体4配体(toll-like receptor 4 ligand,TLR4),可激活TLR4/MyD88依赖性途径,该通路介导了上皮性卵巢癌的化学耐药性和肿瘤进展。苍术内酯I是一种新型TLR4拮抗剂,通过干扰紫杉醇与人白细胞膜TLR4的结合,来抑制TLR4信号传导。Huang等[62]研究发现苍术内酯-I可以减弱紫杉醇诱导的IL-6、VEGF和存活蛋白的蛋白表达,并增强MyD88(+)EOC人卵巢癌细胞的早期凋亡和生长抑制;苍术内酯I被发现更加亲和人髓样分化蛋白2(myeloid differentiation 2,MD-2)的疏水囊,并通过对接模拟与紫杉醇的结合位点部分重叠,这表明苍术内酯-I可能阻断MyD88(+)EOC细胞中MD-2介导的TLR4/MyD88依赖性紫杉醇信号传导。因此,苍术内酯-I可以通过阻断MD-2介导的TLR4/MyD88信号传导,显著提高MyD88(+)EOC细胞对紫杉醇的反应。 结缔组织生长因子(connective Tissue Growth Factor,CTGF)是一种多功能信号调节剂,可通过调节细胞增殖、迁移、侵袭、耐药性和EMT来促进癌症的发生、进展和转移。CTGF还参与大多数节点的肿瘤微环境,包括血管生成、炎症和肿瘤相关成纤维细胞(cancer-associated fibroblasts,CAFs)激活[63]。Wang等[64]研究发现,苍术内酯-I可以下调三阴性乳腺癌细胞中CTGF的表达和分泌。除了通过CTGF抑制三阴性乳腺癌细胞迁移外,苍术内酯-I还下调了成纤维细胞中CTGF的表达,降低了乳腺癌细胞将成纤维细胞转化为CAFs的能力,从而增加了三阴性乳腺癌细胞对紫杉醇的敏感性。在小鼠肿瘤模型中,发现苍术内酯-I治疗可以增强紫杉醇对肿瘤的化疗作用,减少肿瘤向肺和肝的转移。在用苍术内酯-I与紫杉醇联合治疗的小鼠中,源自接种肿瘤的原代培养的成纤维细胞表达相对较低水平的CAFs标志物。 研究表明了苍术内酯-I可以通过阻断CTGF表达和成纤维细胞活化来使三阴性乳腺癌细胞对紫杉醇敏感,还可以通过阻断MD-2介导的TLR4/MyD88信号传导,显著提高肿瘤细胞对紫杉醇的反应并。这些机制有助于未来研究以确定苍术内酯I在临床环境中的价值。对苍术化学成分联合治疗归纳总结,见表3。 图片 4 结语与展望 苍术中含有多种抗肿瘤成分,其中多为倍半萜类成分,如苍术酮、苍术素和苍术内酯等,这些成分多是通过调控PI3K/Akt/mTOR通路来发挥抗肿瘤的作用,但作用靶点与方式却各不相同。例如苍术内酯主要通过降低Akt的磷酸化水平、上调Bax和Bad蛋白表达、增加脂质磷酸酶(PTEN)活性来抑制该通路进而诱导肿瘤细胞凋亡[20];β-桉叶醇能通过激活p27抑制cyclinD1和CDK4蛋白表达最终导致细胞周期停滞于G1期[19]。这些成分通过多途径、多靶点影响肿瘤细胞的生存、运动、代谢和迁移进而共同发挥抗肿瘤作用。正因为其作用机制的不同,使其各有效成分对不同肿瘤的作用具有一定特异性。因此苍术抗肿瘤活性成分联合化疗药物减副增效在科学研究及临床用药时可根据其作用机制进行选择。目前关于苍术化合物对肿瘤细胞的研究还存在一些不足之处,如缺乏对不同肿瘤细胞类型和不同剂量的系统比较、缺乏对苍术化合物与其他药物或放化疗的协同作用的评价,以及缺乏对苍术化合物在体内代谢和药效学的深入分析等。 因此,今后还需要加强对苍术化合物抗肿瘤作用的基础和临床研究。后续可以根据苍术有效成分的抗肿瘤作用机制,筛选出具有最强抗肿瘤活性和最低毒性的化合物,作为候选药物进行进一步的优化和改造,提高其药效和安全性;分析苍术中有效成分的药代动力学特征,研究其在体内的吸收、分布、代谢和排泄等过程,确定其最佳的给药途径、剂量和方案,减少其不良反应和药物相互作用;根据苍术中有效成分的药效学特征,研究其对不同类型、分期和分子标志物的肿瘤细胞的作用差异,确定其最适合的治疗对象和指标,提高其个体化和精准化的治疗效果;根据苍术有效成分的协同增效或拮抗作用,探索其与其他抗癌药物或放化疗的联合应用,实现其对肿瘤细胞的多靶点、多途径和多机制的综合干预,增强其抗肿瘤效能和克服肿瘤耐药性,以期为开发新型的抗肿瘤药物提供更多的选择和可能性。 苍术与化疗药物的联合应用被广泛关注和探索。作为苍术的主要成分,现有研究已表明倍半萜类具有显著的抗肿瘤活性,其与化疗药物的联合临床用药有着巨大的潜力。但倍半萜类化合物分子结构中含有多个疏水基团,导致它们的极性较低,难以与水分子形成氢键或静电相互作用,在水中的溶解度小、生物利用度低。随着现代药物研究技术的现代化和多学科的交叉融合,这些问题也可以通过引入基团、采用纳米技术制备纳米载体、采用共晶技术制备倍半萜类化合物的共晶体等方式来提高其水溶性,进而增强其生物利用度。这些技术在药物化学领域已比较成熟,也已逐步应用于临床药物的开发。例如,抗疟活性药物青蒿素同样具有水溶性差应用困难的问题,通过引入羧酸基团,显著提高了其水溶性和生物利用度[65-66]; 此外,共晶体可以改变倍半萜类化合物的晶型和晶格参数,从而降低其结晶度和熔点,增加其自由能和溶解度[67]。苍术内酯也可通过与尼可替尼(一种具有较高水溶性的抗肿瘤药物)制备共晶体,可以显著提高其水溶性。因此,苍术抗肿瘤有效成分和化疗药物的联合用药在临床环境中的开发和应用具有很高的研究价值。 苍术作为中医临床常用的化湿药。其药性辛、苦、温,归脾、胃、肝经,其苦温燥湿,可以去湿浊、辛温健脾以和脾胃,多用于湿
肿瘤是指体内细胞的异常增生,可以是良性的或恶性的。良性肿瘤(例如息肉)生长缓慢且通常局限在一个区域,不会侵犯周围组织或扩散到其他部位。恶性肿瘤(即癌症)具有侵袭性,可以快速生长并通过血液或淋巴系统扩散到其他身体部位,形成远处转移。癌症是一种严重威胁人类健康和生命的疾病,2020年全球有1 930万新增癌症病例和1 000万癌症死亡病例,且我国癌症发病率和死亡率均位居全球第一[1]。最常见的癌症类型是乳腺癌、肺癌、结直肠癌和前列腺癌。因此,寻找新的抗肿瘤药物,阐明抗肿瘤药物的分子机制,是解决当前临床肿瘤治疗难点的有效策略。中药具有多种有效成分,因其不良反应低、多靶点、多通路等优点,已成为抗肿瘤药物开发的重要来源和研究热点[2]。目前,常规的肿瘤症治疗方法为手术、放射治疗和化学治疗等,但这些方法往往伴随着较大的不良反应和毒性,而且对某些难治性或复发性肿瘤效果不佳[3]。因此,寻找有效、低毒的抗肿瘤药物是当前临床研究的重要方向。 苍术是一种常用的中药材,分为茅苍术Atractylodes lancea (Thunb.) DC.和北苍术A. chinensis (DC.) Koidz.,分别来源于菊科植物茅苍术或北苍术的干燥根茎。苍术具有燥湿健脾、祛风散寒的功效,在《神农本草经》中列为上品[4]。近年来,苍术在抗微生物、抗炎、抗肿瘤、免疫调节、调节消化系统、心血管系统和神经系统等方面的药理作用受到了广泛关注。苍术中含有挥发油、多糖、倍半萜类、聚乙炔类等[5]多种化学成分。其中一些成分已经被证实具有抑制或杀伤多种肿瘤细胞的能力,其作用机制涉及诱导凋亡、抑制增殖、迁移、侵袭和转移,以及调控免疫功能等方面[6]。然而,苍术中的抗肿瘤活性成分及其作用机制尚未完全明确,需要进一步深入地探索和验证。本文通过整理国内外研究文献,对苍术活性成分、苍术与其他药物联合抗肿瘤及其分子机制进行总结,探讨苍术在抗肿瘤方面的应用规律和思路,为苍术资源的开发利用以及抗肿瘤临床疗法的研究提供理论参考。 1 苍术主要化学成分 茅苍术与北苍术化学成分相似,药理作用也较为相似,目前已从苍术中分离出多种化学成分,主要含有包括萜类、聚乙烯炔类、有机酸类、糖苷类化合物等[7-8]。苍术主要抗肿瘤化学成分,见图1。茅苍术与北苍术中主要化学成分如表1所示。 图片 图片 2 苍术的抗肿瘤机制 苍术中含有苍术内酯Ⅰ、Ⅱ、Ⅲ、苍术酮、β-桉叶醇和苍术素等有效成分,这些成分不仅可以抗炎、抗氧化、抗菌、保肝、降血糖,还可以抗肿瘤[14-15]。近年来,苍术及其有效成分对肿瘤的抑制作用受到了广泛的关注。研究发现,苍术有效成分对多种肿瘤细胞都有抑制作用,可以通过多种途径和机制影响肿瘤细胞的生长、迁移、侵袭和血管生成,诱导肿瘤细胞的凋亡和自噬,调节肿瘤微环境和免疫系统。 2.1 抑制肿瘤细胞增殖 肿瘤是由于细胞增殖失控而形成的一种疾病[16]。细胞周期是细胞增殖的基本过程,由细胞周期蛋白(cyclin,CCN)和细胞周期蛋白依赖性激酶(cyclin-dependent kinase,CDK)复合物共同调控[17]。干预细胞周期是抑制肿瘤发展的有效策略之一[18]。Kotawong等[19]发现,苍术中的苍术素、苍术内酯I和β-桉叶醇等有效成分可以通过影响肿瘤细胞周期的不同阶段来抑制肿瘤细胞的增殖。这些成分可以通过抑制磷脂酰肌醇-3-羟激酶(phosphatidylinositol 3-hydroxy kinase,PI3K)、磷酸化蛋白激酶B(protein kinase B,AKT)和哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路来诱导肿瘤细胞在G1期停滞;Yu等[20]发现苍术内酯I通过上调周期蛋白依赖性激酶抑制剂1A(cyclin-dependent kinase inhibitor 1A,p21)和下调cyclinB1、CDK1和细胞分裂周期25C蛋白(cell division cyclin25,Cdc25c)等关键分子来抑制肿瘤细胞在G2/M期的进入,在动物模型中,苍术内酯I可以显著抑制膀胱癌的生长,且无明显不良反应。Zhang等[21]实验发现苍术内酯Ⅱ可以通过改变结直肠癌细胞内的蛋白表达从而抑制结直肠癌细胞的增殖和活性,并且还显著增强了结直肠癌细胞的化疗敏感性。Pongsakorn等[22]发现,苍术提取物可以通过抑制细胞外信号调节激酶信号级联(ERK-signaling cascade,ERK)信号通路来抑制胆管癌细胞的增殖。ERK信号通路是一种重要的细胞内信号转导机制,参与调节细胞生长、分化和凋亡等过程。苍术提取物可以下调ERK及其下游分子的表达,从而抑制胆管癌细胞的生长和增殖,不同类型的胆管癌细胞对苍术提取物的敏感度不同,其中人胆管HuCCT-1癌细胞最为敏感。 2.2 诱导肿瘤细胞凋亡 细胞凋亡是一种程序性细胞死亡形式,它通过限制细胞的增殖和分化来维持组织稳态或去除潜在的有害细胞[23]。目前已知的细胞凋亡途径主要有3种,即外源性途径(死亡受体介导)、内源性途径(线粒体介导)和内质网途径。其中,线粒体途径是最重要的一种,它涉及线粒体外膜透化(outer mitochondrial membrane,MOMP)、细胞色素C释放和半胱天冬酶(cysteine aspartic acid protease,Caspase)激活[24]。多项研究发现,苍术酮可以通过降低线粒体膜电位、提高活性氧水平、抑制B细胞淋巴瘤-2基因(B-cell lymphoma-2,Bcl-2)表达、促进BCL2-相关X蛋白(BCL2-associated X protein,Bax)裂解和Caspase-3表达[25],以及下调PI3K/AKT/mTOR信号通路来诱导肿瘤细胞凋亡[26]。Narahara等[27]研究表明,β-桉叶醇和苍术内酯Ⅲ[27]可以通过增加Caspase-3、Caspase-8、Caspase-9和Bax等凋亡相关蛋白的表达、下调Bcl-2表达、释放细胞色素C和降低线粒体膜电位来诱导胆管癌细胞凋亡。此外,Li等[28]使用β-桉叶醇处理的白血病HL60细胞,发现β-桉叶醇可以通过激活c-JunN端激酶(c-Jun N-terminal kinase,JNK)丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号通路来诱导白血病HL60细胞凋亡。Li等[29]研究发现,苍术素可以通过降低Bcl-2表达、激活p53肿瘤蛋白(p53 tumor protein,p53)、Bax和Caspase-3、-8、-9等凋亡因子来诱导人乳腺癌MCF-7细胞凋亡,并表现出浓度依赖的毒性效应。Li等[30]研究表明,苍术内酯I和苍术内酯Ⅱ[31]可以通过与对两面针激酶2(Janus kinase 2,JAK2)直接相互作用而负调节信号传导及转录激活因子3(signal transducer and activator of transcription 3,STAT3)磷酸化,从而抑制其活化,进而导致糖酵解的抑制和结肠、直肠癌细胞凋亡的诱导。 2.3 抑制肿瘤细胞转移 肿瘤细胞转移是指肿瘤细胞通过血液循环从原发部位转移到其他部位的过程,这是癌症治疗的难点,也是癌症死亡的主要原因[32]。上皮间质转化(epithelial-mesenchymal transition,EMT)是一种与癌症发生相关的细胞程序,它使癌细胞具有移动性、侵袭性和抗凋亡能力,从而促进转移。苍术的一些活性成分具有抑制肿瘤细胞转移的潜在作用,其机制可能涉及对EMT的调控[33]。Acharya等[34]研究发现,β-桉叶醇可以改变EMT相关标志物的表达,从而抑制结肠癌细胞的增殖、迁移和侵袭。同时它还可以影响PI3K、AKT、p38丝氨酸/苏氨酸蛋白激酶(p38 mitogen-activated protein kinase,p38MAPK)信号通路,以及肺癌细胞中的活性氧水平,从而降低癌细胞的黏附和迁移能力[35]。麦静愔等[36]发现苍术酮可以通过抑制EMT过程等途径抑制肿瘤细胞的迁移和侵袭能力,此外,苍术酮还可以通过下调基质金属蛋白酶(matrix metalloproteinase,MMP)的表达从而抑制肿瘤细胞的迁移和侵袭能力。MMP是一类能够降解细胞外基质(extracellular matrix,ECM)的锌依赖性内肽酶,在癌症进展中的作用与它们参与ECM降解以及黏附和细胞骨架蛋白、生长因子、趋化因子的调节和加工有关[37]。且有动物实验表明,苍术酮可以明显抑制肝癌生长,没有明显的毒性。Zhong等[38]在观察了苍术多糖在U-2 OS人骨肉瘤细胞中对内皮细胞选择素(endothelial cell selectin,E-Selectin)和路易斯X三糖(Lewis-X Trisaccharide,LacCer Lex)的影响,发现苍术多糖可通过降低U-2 OS细胞上的E-Selectin抑制U-2 OS细胞对人脐静脉内皮细胞HUVECs的黏附、迁移和侵袭。肿瘤相关巨噬细胞(tumor-associated macrophages,TAMs)在促进肿瘤转移中发挥重要作用,Zhang等[39]发现苍术内酯II可以有效抑制肿瘤细胞极化,从而抑制肺癌细胞在体内和体外的转移。铁死亡是一种新的细胞死亡模式,其特征是铁过载导致脂质过氧化而导致膜损伤,过度的铁死亡会影响肿瘤的转移,从而抑制肿瘤的进展[40]。He等[41]发现,苍术素可通过抑制谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)和铁蛋白轻链(ferritin light chain,FTL)的表达,以及上调酰基辅酶A合成酶长链家族成员4(acyl-CoA synthetase long-chain family member 4,ACSL4)和转铁蛋白受体(transferrin receptor,TFR1)的表达来诱导肝癌HCCM细胞的铁死亡。 2.4 诱导肿瘤细胞自噬 细胞自噬是一种分解代谢通路,能清除不必要的或功能失调的细胞成分并回收代谢底物[42]。目前已知有3种主要的细胞死亡方式:细胞凋亡(Ⅰ型)、自噬性细胞死亡(Ⅱ型)和坏死(Ⅲ型)。自噬性细胞死亡是指自噬过程中产生的自噬体过多或过大,导致细胞质溶解和细胞死亡。自噬体是由双层膜包裹的囊泡,内含被降解的细胞器和蛋白质。微管相关蛋白1轻链3(microtubule-associated protein 1 light chain 3,LC3)是自噬体形成的关键标志物,它以微管相关蛋白1A/1B-轻链3(microtubule-associated protein 1 light chain 3,LC3-I)和微管相关蛋白1轻链3的脂化形式(lipidated form of microtubule-associated protein 1 light chain 3,LC3-Ⅱ)2种形式存在,LC3-Ⅰ转化为LC3-Ⅱ是自噬体形成的必要步骤[43-44]。Li等[29]使用苍术素处理乳腺癌MCF-7细胞时发现,苍术素可以增加了LC3Ⅰ向其脂化形式的LC3Ⅱ的转化,并增加了苄氯素1(beclin-1,BECN1)的表达,下调了人乳腺癌MCF-7细胞中的p62蛋白(p62 protein,p62)表达,改变凋亡和自噬相关生物标志物。Acharya等[45]研究发现,苍术素通过调节PI3K、AKT、mTOR、p38MAPK信号通路的活性,可以诱导胆管癌HuCCT-1细胞发生自噬,并抑制其生长、迁移和侵袭,SB202190(p38MAPK诱导剂)和3-MA(p38MAPK抑制剂)分别显著增加和降低苍术素诱导的自噬速率。 2.5 抑制肿瘤血管生成 血管生成本身不会导致恶性肿瘤的形成,但可以为肿瘤的生长和转移提供条件。肿瘤在发展到一定阶段后,需要依赖新生血管来满足其对氧气和营养的增加的需求,以及排除代谢废物,因此,抑制血管生成是一种有效的抗肿瘤策略[46]。血红素加氧酶1(heme oxygenase 1,HO-1)是一种在肿瘤组织中高表达的酶,它可以促进肿瘤的血管生成和抗氧化应激,为肿瘤细胞提供生存优势。因此,抑制HO-1的表达或活性是治疗肿瘤的另一种有效策略之一。Mathema等[47]研究发现,苍术素可以抑制胆管癌CL6肿瘤细胞的集落形成和伤口愈合能力,其机制与抑制HO-1的表达、下调信号转导及转录激活蛋白1/3(signal transducer and activator of transcription 1/3,STAT1/3)和核因子κB(nuclear factor kappa-B,NF-κB)的信号通路有关。β-桉叶醇也具有抑制胆管癌细胞中HO-1的表达的能力,其机制与浓度依赖性地抑制STAT1/3和NF-κB信号通路有关[48]。β-桉叶醇还可以通过抑制生长因子信号通路中的环磷腺苷效应元件结合蛋白(cyclic-AMP response binding protein,CREB)激活来阻断血管生成,从而抑制肿瘤的发展[49]。Tsuneki等[50]有动物实验表明,β-桉叶醇可以通过激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)来刺激大鼠嗜铬细胞瘤细胞中的神经突生长,且β-桉叶醇还表现出了体外和体内的抗血管生成活性,其阻断了由碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)或血管内皮生长因子(vascular endothelial growth factor,VEGF)诱导的人脐静脉内皮细胞(human umbilical vein endothelial cell,HUVEC)中CREB蛋白的磷酸化,从而抑制bFGF刺激的HUVEC迁移和HUVEC在基质胶中的管形成。同时,它还能显著降低小鼠皮下植入的Matrigel栓塞和小鼠佐剂诱导的肉芽肿中的血管生成[51]。 2.6 免疫调节作用 随着肿瘤的发生和发展,或在接受化疗、放疗等治疗的过程中,肿瘤患者机体免疫力的显著下降。因此,调节或刺激机体免疫能力,可能是一种有效的主动抗癌策略。免疫治疗作为一种新型的抗癌手段,已经引起了广泛的关注和研究[52]。巨噬细胞是机体内重要的免疫细胞,在机体免疫中发挥着重要的作用[53]。Qin等[54]从苍术中分离得到两种多糖成分:中性多糖和酸性多糖。研究表明,酸性多糖能够显著地刺激小鼠单核巨噬细胞白血病细胞(RAW264.7)细胞的增殖、吞噬能力、NO产生和细胞因子分泌,并且呈现出剂量相关性,而中性多糖则相对较弱。此外,中性多糖和酸性多糖均能够激活淋巴结Peyers patch细胞中的T细胞,并促进集落刺激因子的产生。而酸性多糖也表现出比中性多糖更好的肠道免疫调节活性。吲哚胺-2,3-二氧化酶(indoleamine 2,3-dioxygenase,IDO)是一种通过犬嘌呤途径氧化分解色氨酸的限速酶,是抗肿瘤免疫治疗中小分子药物开发的潜在目标。IDO可在肿瘤微环境中通过与许多肿瘤相关的自发炎症和T细胞激活而被诱导。Liu等[55]研究发现,苍术内酯Ⅰ可以通过下调Toll样受体4/髓样分化蛋白2复合物(toll-like receptor 4/myeloid differentiation 2 complex,TLR4/MD-2)的表达,抑制人卵巢癌细胞(EOCSKOV3)中髓样分化主要反应蛋白88(myeloid differentiation primary response protein 88,MyD88)、NF-κB、Akt和IDO1的信号通路的活化,从而减少白细胞介素-6(interleukin-6,IL-6)、转化生长因子-β1(transforming growth factor beta 1,TGF-β1)、VEGF和白细胞介素-17A(interleukin-17A,IL-17A)等促进肿瘤免疫逃逸的因子的分泌。同时,还可以降低调节性T细胞(Treg细胞)在肿瘤微环境中的比例,改善T淋巴细胞受到EOCSKOV3细胞上清液抑制而导致的增殖反应降低和抗肿瘤细胞毒性减弱。Liu等[56]研究发现,苍术内酯Ⅲ可以通过直接结合JAK3蛋白,从而抑制γ-干扰素(interferon gamma,IFN-γ)触发的JAK3/STAT3通路,从而达到抑制IDO激活的目的。 苍术抗肿瘤成分的潜在分子机制见图2。对苍术抗肿瘤有效成分及其抗肿瘤作用进行归纳总结,见表2。 图片 图片 3 联合用药 西医治疗肿瘤的常用手段有手术切除、药物化疗和高能射线放疗等,这些手段去除肿瘤西医的治疗方式更为直接,适合前期控制病情,化疗药物虽然能够杀死肿瘤细胞,但同时也伴有严重的副作用,影响患者的生活质量和治疗效果。中药具有不良反应小、安全性高的特点,因此中药与化疗药物的联合应用被广泛关注和探索[57]。 阿帕替尼是全球第一个在晚期胃癌被证实安全有效的小分子抗血管生成靶向药物,也是晚期胃癌标准化疗失败后,明显延长生存期的单药。Zhou等[58] 分析了不同苍术多糖提取方法的影响。比较了热水浸提法、超声浸提法和酶浸提法提取苍术多糖的得率、总糖含量、相对分子质量分布、单糖组成、并测定苍术多糖与阿帕替尼的协同活性。结果发现其中超声浸提法表现出最强的协同作用。这也与超声浸提的苍术多糖相对分子质量小、β-构型高、半乳糖含量高的事实相一致。Srijiwangsa等[59]发现,β-桉叶醇可以通过抑制胆管癌细胞和细胞裂解物中的NAD(P)H醌氧化还原酶1[NAD(P)H quinonedehydrogenase 1,NQO1]的活性和蛋白表达,增强氟尿嘧啶和多柔比星对细胞迁移的细胞毒性活性和抑制活性。Mai等[60]将不同浓度的苍术内酯I、硼替佐米以及硼替佐米+苍术内酯I作用于U266细胞结果研究发现,苍术内酯可以调节JAK2/STAT3通路上的IL-6、JAK2、STAT3等基因表达抑制U266肿瘤细胞的增殖和促进其凋亡并呈剂量依赖性,并能与硼替佐米产生协同作用,当苍术内酯I与硼替佐米联合使用时,可显著增强对U266细胞增殖的抑制作用。 紫杉醇是第一个获得批准的草药衍生化疗药物[61]。并且作为一种已知的Toll受体4配体(toll-like receptor 4 ligand,TLR4),可激活TLR4/MyD88依赖性途径,该通路介导了上皮性卵巢癌的化学耐药性和肿瘤进展。苍术内酯I是一种新型TLR4拮抗剂,通过干扰紫杉醇与人白细胞膜TLR4的结合,来抑制TLR4信号传导。Huang等[62]研究发现苍术内酯-I可以减弱紫杉醇诱导的IL-6、VEGF和存活蛋白的蛋白表达,并增强MyD88(+)EOC人卵巢癌细胞的早期凋亡和生长抑制;苍术内酯I被发现更加亲和人髓样分化蛋白2(myeloid differentiation 2,MD-2)的疏水囊,并通过对接模拟与紫杉醇的结合位点部分重叠,这表明苍术内酯-I可能阻断MyD88(+)EOC细胞中MD-2介导的TLR4/MyD88依赖性紫杉醇信号传导。因此,苍术内酯-I可以通过阻断MD-2介导的TLR4/MyD88信号传导,显著提高MyD88(+)EOC细胞对紫杉醇的反应。 结缔组织生长因子(connective Tissue Growth Factor,CTGF)是一种多功能信号调节剂,可通过调节细胞增殖、迁移、侵袭、耐药性和EMT来促进癌症的发生、进展和转移。CTGF还参与大多数节点的肿瘤微环境,包括血管生成、炎症和肿瘤相关成纤维细胞(cancer-associated fibroblasts,CAFs)激活[63]。Wang等[64]研究发现,苍术内酯-I可以下调三阴性乳腺癌细胞中CTGF的表达和分泌。除了通过CTGF抑制三阴性乳腺癌细胞迁移外,苍术内酯-I还下调了成纤维细胞中CTGF的表达,降低了乳腺癌细胞将成纤维细胞转化为CAFs的能力,从而增加了三阴性乳腺癌细胞对紫杉醇的敏感性。在小鼠肿瘤模型中,发现苍术内酯-I治疗可以增强紫杉醇对肿瘤的化疗作用,减少肿瘤向肺和肝的转移。在用苍术内酯-I与紫杉醇联合治疗的小鼠中,源自接种肿瘤的原代培养的成纤维细胞表达相对较低水平的CAFs标志物。 研究表明了苍术内酯-I可以通过阻断CTGF表达和成纤维细胞活化来使三阴性乳腺癌细胞对紫杉醇敏感,还可以通过阻断MD-2介导的TLR4/MyD88信号传导,显著提高肿瘤细胞对紫杉醇的反应并。这些机制有助于未来研究以确定苍术内酯I在临床环境中的价值。对苍术化学成分联合治疗归纳总结,见表3。 图片 4 结语与展望 苍术中含有多种抗肿瘤成分,其中多为倍半萜类成分,如苍术酮、苍术素和苍术内酯等,这些成分多是通过调控PI3K/Akt/mTOR通路来发挥抗肿瘤的作用,但作用靶点与方式却各不相同。例如苍术内酯主要通过降低Akt的磷酸化水平、上调Bax和Bad蛋白表达、增加脂质磷酸酶(PTEN)活性来抑制该通路进而诱导肿瘤细胞凋亡[20];β-桉叶醇能通过激活p27抑制cyclinD1和CDK4蛋白表达最终导致细胞周期停滞于G1期[19]。这些成分通过多途径、多靶点影响肿瘤细胞的生存、运动、代谢和迁移进而共同发挥抗肿瘤作用。正因为其作用机制的不同,使其各有效成分对不同肿瘤的作用具有一定特异性。因此苍术抗肿瘤活性成分联合化疗药物减副增效在科学研究及临床用药时可根据其作用机制进行选择。目前关于苍术化合物对肿瘤细胞的研究还存在一些不足之处,如缺乏对不同肿瘤细胞类型和不同剂量的系统比较、缺乏对苍术化合物与其他药物或放化疗的协同作用的评价,以及缺乏对苍术化合物在体内代谢和药效学的深入分析等。 因此,今后还需要加强对苍术化合物抗肿瘤作用的基础和临床研究。后续可以根据苍术有效成分的抗肿瘤作用机制,筛选出具有最强抗肿瘤活性和最低毒性的化合物,作为候选药物进行进一步的优化和改造,提高其药效和安全性;分析苍术中有效成分的药代动力学特征,研究其在体内的吸收、分布、代谢和排泄等过程,确定其最佳的给药途径、剂量和方案,减少其不良反应和药物相互作用;根据苍术中有效成分的药效学特征,研究其对不同类型、分期和分子标志物的肿瘤细胞的作用差异,确定其最适合的治疗对象和指标,提高其个体化和精准化的治疗效果;根据苍术有效成分的协同增效或拮抗作用,探索其与其他抗癌药物或放化疗的联合应用,实现其对肿瘤细胞的多靶点、多途径和多机制的综合干预,增强其抗肿瘤效能和克服肿瘤耐药性,以期为开发新型的抗肿瘤药物提供更多的选择和可能性。 苍术与化疗药物的联合应用被广泛关注和探索。作为苍术的主要成分,现有研究已表明倍半萜类具有显著的抗肿瘤活性,其与化疗药物的联合临床用药有着巨大的潜力。但倍半萜类化合物分子结构中含有多个疏水基团,导致它们的极性较低,难以与水分子形成氢键或静电相互作用,在水中的溶解度小、生物利用度低。随着现代药物研究技术的现代化和多学科的交叉融合,这些问题也可以通过引入基团、采用纳米技术制备纳米载体、采用共晶技术制备倍半萜类化合物的共晶体等方式来提高其水溶性,进而增强其生物利用度。这些技术在药物化学领域已比较成熟,也已逐步应用于临床药物的开发。例如,抗疟活性药物青蒿素同样具有水溶性差应用困难的问题,通过引入羧酸基团,显著提高了其水溶性和生物利用度[65-66]; 此外,共晶体可以改变倍半萜类化合物的晶型和晶格参数,从而降低其结晶度和熔点,增加其自由能和溶解度[67]。苍术内酯也可通过与尼可替尼(一种具有较高水溶性的抗肿瘤药物)制备共晶体,可以显著提高其水溶性。因此,苍术抗肿瘤有效成分和化疗药物的联合用药在临床环境中的开发和应用具有很高的研究价值。 苍术作为中医临床常用的化湿药。其药性辛、苦、温,归脾、胃、肝经,其苦温燥湿,可以去湿浊、辛温健脾以和脾胃,多用

片姜黄提取物14种挥发油的抗肿瘤活性研究试验 从国内外对姜黄挥发油的药理活性的研究报道发现,姜黄挥发油具有显著的抗肿瘤、抑制细胞增殖的作用。他们可通过直接杀伤肿瘤细胞、抑制肿瘤细胞生长或转移、诱导肿瘤细胞凋亡或诱导肿瘤细胞分化使其逆转、增强和刺激机体免疫功能等多种方式起到抗肿瘤作用。 本研究采用体外方法评价了从片姜黄中提取分离得到的14种挥发油单体化合物在体外对HL-60(人白血病细胞)、Du145(人前列腺癌细胞)、Hep-3B(人肝癌细胞)、U87(人胶质瘤细胞)的细胞毒活性。实验结果表明,部分受试化合物显示出不同强度的抗肿瘤活性。挥发油单体结构式如下:http://ng1.17img.cn/bbsfiles/images/2014/11/201411281001_525011_2188679_3.jpghttp://ng1.17img.cn/bbsfiles/images/2014/11/201411281001_525012_2188679_3.jpg实验部分1实验材料1.1受试品14个单体化合物(化合物1-14)由合作单位离得到并进行结构鉴定,阳性对照购自药店。1.2 实验细胞株HL-60(人白血病细胞),Du145(人前列腺癌细胞),Hep-3B(人肝癌细胞),U87(人胶质瘤细胞),购自ATCC2实验试剂 RPMI1640培养基:Gibco 胎牛血清:天津市灏洋生物制品有限公司甲基偶氮唑盐(MTT):美国Sigma二甲基亚砜(DMSO),NaCl,KCl,KH2PO3,Na2HPO3,NaHCO3:沈阳化学试剂厂酶标仪:奥地利TECAN 96孔细胞培养板:Costar公司3.试验方法3.1药物的处理[fon
不知各位老大对诱导含量的定义有了解不,以及如何使用诱导含量,谢谢
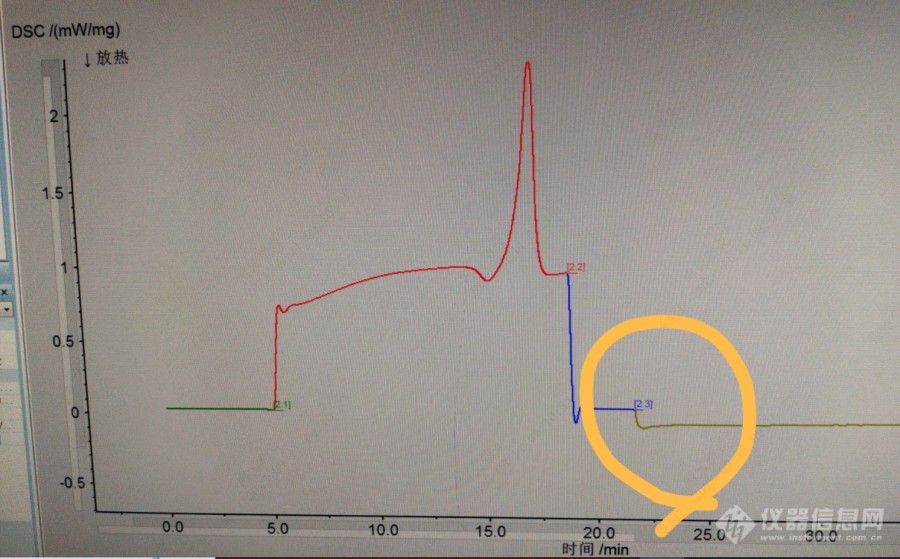
氧化诱导时间的测试一定要过样品的熔点吗?1、我的样品只要转氧就会立即氧化;即使我将温度设定稍低于熔点了还是有这样的趋势;材料是尼龙,所以氧化诱导时间一定要过材料的熔点测试才有意义吗?标准里好像也没有这样的规定。2、如果温度太低呢,氧化诱导时间可能会很长,难道尼龙就不适合做氧化诱导时间?氧化诱导温度我做过,没什么问题,在320℃左右。[img=,690,428]https://ng1.17img.cn/bbsfiles/images/2023/09/202309110914235323_5961_3929349_3.jpg!w690x428.jpg[/img]
各位前辈 为什么我做的所有氧化诱导时间的分析测试结果都在1分钟左右啊 氧化诱导期的影响因素都有哪些啊
最近在开发GB/T 19466.6-2009氧化诱导期的测试方法,实验室内用LDPE测的氧化诱导期温度偏差大了点,想改善重复性。排除制样后,氧气的纯度没确认,但在空气中的重复性也不好,还可能是什么原因呢?
各国争相发展的重点项目 iPS技术,即诱导性多能干细胞技术,是一种将成体成熟、分化的体细胞重编程获得类似胚胎干细胞的新兴技术。2007年11月美国和日本科学家分别独立宣布可将人类皮肤细胞转化为iPS细胞。这一发现被《自然》和《科学》杂志分别评为2007年第一和第二大科学进展。之后,iPS细胞研究迅猛发展,不同的国家和实验室纷纷报道了多种方法建立的iPS细胞系。就连世界第一只体细胞克隆动物多利羊的培育者伊恩·威尔莫特也宣布放弃人类胚胎干细胞克隆研究,转而进行 iPS 细胞研究,因为他认为这种细胞比胚胎干细胞更具潜在优势。 我国连续多年将干细胞研究列入“863”、“973”、国家自然基金重点项目。国务院2006年发布的《国家中长期科学和技术发展规划纲要(2006-2020年)》中,干细胞作为五项生物技术之一成为未来15年我国前沿技术的重点研究领域。 致瘤风险浮出水面 Yamanaka研究组在《自然·生物技术》上发表的文章显示,用iPS细胞诱导的神经干细胞,即使不含c-Myc(曾被认为是导致肿瘤的主要原因),在植入NOD/SCID免疫缺陷小鼠后仍有很强的致瘤性,甚至高于胚胎干细胞。 他们共研究了36个iPS细胞克隆,在诱导方式上,有些诱导剂配方中含有c-Myc基因,有些没有,因此具有较好的代表性。同时他们选择了3株胚胎干细胞作为对照。在45周的观察中,移植胚胎干细胞来源神经干细胞的34只小鼠有4只长出肿瘤。在100只移植胚胎成纤维细胞来源的iPS神经干细胞小鼠中34只发现肿瘤,概率和胚胎干细胞相当。在55只移植成人成纤维细胞来源的iPS神经干细胞小鼠中46只发现肿瘤,概率远高于胚胎干细胞。在36只移植肝细胞来源的iPS神经干细胞小鼠中10只发现肿瘤,概率高于胚胎干细胞。8只移植胃上皮细胞来源的iPS神经干细胞小鼠中未发现肿瘤。病理学检查证实肿瘤均为畸胎瘤,部分为恶性畸胎瘤。 研究还发现,以前认为致瘤性很强的c-Myc在去掉后并没有减少iPS神经干细胞的致瘤性,相反以前认为没有致瘤性的Nanog基因却可以明显增强iPS神经干细胞的致瘤性。 这次试验的另一个意外结果是并未发现在生成的肿瘤细胞中有c-Myc或其他基因的激活。以前的观点认为,转入的癌基因是iPS致瘤性的基础,只要在iPS细胞诱导成功后通过各种方法去除已完成使命的癌基因即可使iPS细胞免于致瘤性。这次试验的结果无疑给这些想法留下了阴影,而且使iPS致瘤的机制更加扑朔迷离。
[align=center]抗肿瘤单克隆抗体药物的研究进展[/align][align=center] [/align][align=center]摘 要[/align][align=center] [/align] 通过淋巴细胞杂交瘤技术或基因工程技术制备单克隆抗体药物,已经成为生物制药领域的一个重要方面,特别是对抗肿瘤单克隆抗体药物的研究已获得了重要进展。多年来,许多研究证实了抗肿瘤单克隆抗体药物的作用,为其应用于肿瘤治疗提供了重要依据。这类药物的特异性强,疗效显著。本文主要就近年来抗肿瘤单克隆抗体药物的研究进展进行了综述,并对抗肿瘤单克隆抗体药物的发展前景进行了展望。[align=left] [/align][align=left]关键词:抗肿瘤;单克隆抗体;研究进展[/align][align=center] [/align][align=center] [/align][b]一 引言[/b]抗肿瘤单抗药物因与烷化剂、抗代谢药、抗肿瘤抗生素、铂类配合物、植物药等抗肿瘤药物相比,具有高效价、高特异性、血清交叉反应少等特点与优点,在肿瘤治疗中起着不可替代的作用。单抗药物是当前生物技术药物领域甚为活跃的部分。针对特定的分子靶点(抗原),单抗有高度特异性。针对各种不同的抗原,可以制备为数众多的、各不相同的单抗;因此,作为药物来源,单抗又具有高度多样性。由于其特异性和多样性,研制单抗药物有巨大的潜力。单克隆抗体药物治疗恶性瘤主要机制有两种[sup][/sup]:一是利用单克隆抗体本身来阻断癌细胞生长的信号,单克隆抗体在癌细胞膜外与生长因子竞争结合受体,阻断信号传递过程,从而阻止癌细胞的生长和扩散,诱导细胞凋亡或者间接激活宿主的抗肿瘤免疫反应;二是利用单克隆抗体作为药物载体的靶向治疗,如将有细胞毒性的药物或有放射性的药物靶向性的运送到肿瘤细胞,从而杀伤肿瘤细胞。目前,国际上与肿瘤治疗相关的抗体研究主要集中在将抗体与耦联物作用后直接杀伤肿瘤细胞,利用抗体促进肿瘤细胞凋亡和抑制肿瘤血管生成等方面。此外,研究表明静脉内注射抗肿瘤单抗,在肿瘤部位的浓度较高,显示特异性定位;单抗与药物的偶联物通常仍保留原来单抗的分布特征,在靶肿瘤的浓度较高[sup][/sup]。[align=center]二 单克隆抗体药物作用靶点[/align]特定受体或特定的基因表达蛋白可能作为单抗药物的靶点。Rituxan是以B细胞的CD20分子作为靶点的人鼠嵌合抗体,对非霍奇金氏B细胞淋巴瘤有疗效,是第一个获美国FDA批准用于治疗恶性肿瘤的单抗。Herceptin是抗HER-2/neu癌基因编码蛋白的单抗,临床研究对乳腺癌有效,与化疗药物联合有更显著的疗效。Mylotarg是由抗CD33单抗与calicheamicin构成的偶联物,已获批准用于治疗急性复发性髓性白血病[sup][/sup]。表皮细胞生长因子受体(EGFr)在人的鳞癌、乳腺癌和脑胶质瘤等均有较高的表达。有报道,抗EGFr单抗与长春碱衍生物的偶联物在裸鼠体内试验,显示良好的抗癌效果。抗EGFr的人鼠嵌合抗体已进入临床研究。血管内皮生长因子(VEGF)在血管生成中有重要作用。据报道,抗VEGF的中和性单抗具有广谱的抗肿瘤作用,对移植于裸鼠的人体癌瘤有显著疗效[sup][/sup]。[b]三 单抗诱发肿瘤细胞凋亡[/b][align=left] 3.1 通过免疫细胞表面抗原的交联作用而诱导恶性肿瘤细胞的凋亡[/align]用于治疗血液系统恶性肿瘤的单克隆抗体药物大多是通过免疫细胞表面抗原的交联作用诱导恶性肿瘤细胞凋亡而起作用的,如目前用的抗-CD20的单克隆抗体——美罗华。其单克隆抗体的作用机制是通过诱导CD20分子在B细胞膜上的脂筏区聚集,再在一系列激酶的作用下使脂筏信号传导区域的CD20分子亲和性增强,从而形成CD20交联形式;交联的CD20分子启动了细胞内凋亡信号的传导通路,使线粒体释放出细胞色素C,激活下游的caspase级联反应,最终导致细胞凋亡[sup][/sup]。3.2 作用于恶性肿瘤细胞膜上的生长因子及其受体而诱导细胞凋亡许多生长因子及其受体通过作用于细胞的存活途径、刺激细胞的有丝分裂、促进细胞的生长增殖来阻止细胞凋亡。与正常细胞中生长因子信号传导的严格调控相比,肿瘤细胞中的失控则导致细胞的恶性增殖,从而使恶性细胞获得“永生”。单克隆抗体通过作用于恶性肿瘤细胞膜上的生长因子及其受体可阻断存活信号传导通路,从而导致其凋亡,同时还能对化疗和放疗有正协同作用。目前主要集中在对血管内皮生长因子(VEGF)及其受体、表皮生长因子受体(EGFR)等的研究。美国FDA于2006年批准了第一个用于治疗头颈部鳞状细胞癌的单克隆抗体药物——Cetuximab,它为一种IgG1单克隆抗体,主要通过干扰癌细胞表面EGFR的生长,从而减少癌细胞进入正常组织的概率,控制癌细胞的转移,达到抗癌目的[sup][/sup]。最初想到制备针对恶性肿瘤凋亡相关分子的单克隆抗体药物时,虽然从理论上来说无疑是给人们注入了一针兴奋剂,但在实际应用中则并不然,所以在通过单克隆抗体药物诱导恶性肿瘤细胞凋亡的研究和治疗中,还有待进一步开发新的、更经济、更有效地药物。[b]四 单克隆抗体耦联物[/b]4.1 抗体与化疗药物耦联目前,国内外研究较多的与单克隆抗体耦联的化学药物有平阳霉素、柔红霉素、丝裂霉素、多柔比星(阿霉素)、顺铂以及长春碱类衍生物等。同时还可以通过脂质体靶向制剂作为化疗药靶向治疗肿瘤,利用脂质体制剂将药物导向靶标进行有选择性地杀伤癌细胞和抑制癌细胞的繁殖,以达到提高疗效和高度定向作用。目前已上市的脂质体有复方氟脲嘧多相脂质体、喜树碱多相脂质体、阿霉素脂质体和紫杉醇脂质体等。4.2 抗体导向酶耦联利用抗体与肿瘤细胞表面抗原的特异性结合,将前体药物的专一性活化酶与单抗耦联,导向输入到靶细胞部分,再注入前体药物,使其在酶的作用下转化为活性药物,进而杀伤肿瘤细胞[sup][/sup]。目前这种用作前体药物的抗癌药有苦杏仁苷、氮芥、鬼臼乙叉苷、阿霉素、丝裂霉素等。而作为活化前体药物的导向酶有碱性磷酸酶、青霉素V或G酰胺酶、羧基酶肽、胸腺嘧啶核苷酶、β葡萄苷酶等。临床研究表明,单抗耦联物对于抗药性肿瘤细胞仍显示较强的杀伤活性。对由于长期使用氨甲蝶呤而出现抗药性的成骨肉瘤细胞,单抗氨甲蝶呤耦联物仍显示较强的杀伤作用。对于具有多药抗药性(MDR)的肿瘤细胞,抗P-170糖蛋白单抗构成的免疫毒素可显示选择性杀伤作用[sup][/sup]。这说明,单抗药物有可能用于克服肿瘤细胞抗药性。[b]五 单克隆抗体靶向药物[/b]单抗靶向药物是利用单抗对肿瘤表面相关抗原或特定的受体特异性识别,从而把药物直接导向肿瘤细胞,提高药物的疗效,降低药物对循环系统及其他部位的毒性。研究表明,单抗靶向药物具有很好的疗效,在免疫偶联物对移植于裸鼠的相应人体肿瘤生长有抑制作用。免疫偶联物与相应的游离物比较,具有更高更好的疗效和较低的细胞毒性[sup][/sup]。单克隆抗体体积小,能更有效地透入肿瘤;分子小、消除快、累积毒性小;所携带的弹头脱离后,可较快被清除 循环中免疫靶向结合物对靶细胞的竞争作用小;半衰期短;穿透性好;能穿过血脑屏障[sup][/sup],因而还可以作为新一代靶向载体。与化学药物、毒素、放射性核素、生物因子、基因、分化诱导剂、光敏剂、酶等物质构成单克隆抗体靶向药物,把杀伤肿瘤细胞的活性物质特异的输送到肿瘤部位,利用单抗对肿瘤表面相关抗原或特定的受体特异性识别,从而把药物直接导向肿瘤细胞,提高药物疗效,降低药物对循环系统及其他部位的毒性。近年来,随着医学、药学和生物工程学及技术的进步,临床对肿瘤的根治和对癌细胞的攻击锁定于表皮生长因子和血管内皮生长因子等靶位,使药物治疗的切入点由细胞水平提升到分子和抗体水平,从而提高了肿瘤综合治疗的效果。[align=center]六 人源化单克隆抗体[/align]单克隆抗体是近年竞相开发的品种,自1997年第1个单克隆抗体rituximab通过食品与药物管理局(FDA)批准应用于临床以来,目前已经上市的单克隆抗体靶向药物的疗效令人瞩目,在抗肿瘤、类风湿性关节炎和自身免疫系统缺陷治疗领域得到了有力的推广,其以独特的作用优势,在肿瘤的治疗中不但能够选择性杀伤癌细胞,且在体内表现出特异的分布特性,具有高效、低毒的特点,从而在生物技术产品领域中占据了1/3的市场[sup][/sup]。目前用于治疗肿瘤的单克隆抗体药已有多个,包括伊珠单抗奥加米星、帕尼单抗、曲妥珠单抗等。伊珠单抗奥加米星又名CMC-544,是以人源化抗CD22的抗体伊珠单抗与 CalichDMH偶联形成的ADC药物,用来治疗复发性或难治性B细胞非霍奇金淋巴瘤(B cell-NHL)和急性淋巴细胞白血病(ALL),目前已经进入临床 III期试验[sup][/sup]。帕尼单抗是一种IgG2单克隆抗体完全人源化可以与EGFR高度特异性地结合,进而阻断配体诱导的信号激活,从而抑制肿瘤生长。有临床研究选择既往未治疗过的ⅢB或Ⅳ期非小细胞肺癌患者比较卡铂(AUC=6,每3周),加紫杉醇(200mg/m21次/3周) 联合或不联合帕尼单抗(2.5 mg/m2,1次/周) 化疗的疗效及其安全性。研究结果显示,单纯化疗组与帕尼单抗联合化疗组之间在PFS(5.3个月对比4.2个月、P=0.55)和总生存( Overall survival,OS)(8.0个月对比8.5个月,P =0.81)上均无显著差异。结果提示帕尼单抗联合一线化疗方案可能对晚期非小细胞肺癌无明显疗效[sup][/sup]。曲妥珠单抗是一种抗Her2的单克隆抗体,他可以和肿瘤细胞的HER2/neu特异性地结合,从而阻断细胞内生长信号的转导,同时曲妥珠单抗还可以诱导体内巨噬细胞以及自然杀伤细胞攻击肿瘤细胞,以达到抑制和杀伤肿瘤细胞的目的。比较用或不用曲妥珠单抗联合一线化疗方案用以治疗ⅡB/Ⅲ期HER2/neu阳性的 NSCLC患者差异的两项大型的随机Ⅱ期临床试验,其结果显示两个试验结论相似,曲妥珠单抗不能提高化疗的疗效,但也不加重化疗的不良反应。试验中HER2/neu值为3+的患者对曲妥珠单抗治疗的反应性较好,提示曲妥珠单抗对这一较少见类型的NSCLC效果要更好[sup][/sup]。在临床治疗中使用鼠派生单抗的主要障碍之一是产生人抗鼠抗体(HAMA)反应,通过基因工程技术制备嵌合抗体的I-IPdVIA反应率较鼠源性单抗低,但完全的人源抗体才是单抗药物的发展目标。噬茵体抗体库技术和转基因小鼠技术是制备完全人源单抗的两种方法[sup][/sup]。因此,只有不断地完善单克隆抗体人源化的技术,才能更好地将完全人源化的单克隆抗体用于肿瘤分子靶向治疗中,从而使医学界迈向更高的台阶。[b]七 问题与对策[/b]在限制单克隆抗体临床治疗效果的因素有:(l)循环免疫复合物导致的肝肾功能损害。(2)可溶性肿瘤抗原释放造成的体液中的封闭作用。(3)异种蛋白反应。(4)特异性还不够专一,引起了正常细胞的伤害。(5)天然免疫功能低下(如补体介异的细胞毒,网状内皮系统清除和ADCC作用等)。(6)主要的问题还在这种免疫疗法会导致靶细胞(肿瘤细胞)上抗原的转换。为了解决这些问题,今后的研究应着重:(1)制备对肿瘤抗原有高度特异性的单克隆抗体。(2)选择不易诱导抗原转换的单克隆抗体。(3)研究副作用较少,既安全疗效又高的偶联制剂。单抗(Mab)药物存在的一个最关键问题就是人抗鼠抗体反应(HAMA)。由于用于临床研究的Mab药物一般使用鼠源Mab,这不可避免地会引起HAMA反应,所以尽量避免HAMA反应这一副作用才是Mab药物能否真正适合治疗肿瘤性疾病的重点[sup][/sup]。近些年来,Mab药物的研究主要是向减轻宿主对外源抗体的排斥,促进抗体人源化,改变抗体的氨基酸序列而增加或降低该抗体的生物学效应,加抗体的亲和力,制备双特异性抗体,改造抗体重链恒定区以增强抗体功能,以及寻找新的分子靶点(相对特异的肿瘤抗原)等方向发展[sup][/sup]。Mab药物的不断更新,必将为全球的肿瘤患者带来更大的希望。[align=center]八 总结与展望[/align]目前肿瘤治疗中使用最广泛的仍是化疗以及放射性疗法,其毒副作用较大。随着基因工程技术和DNA重组技术的兴起,利用单克隆抗体治疗肿瘤已经日渐取代副作用较大的传统疗法而成为新的发展趋向。所以,如何研制更多的单克隆抗体以及怎样更好的利用单克隆抗体治疗肿瘤,将成为肿瘤治疗研究中的又一艰巨任务。同时,生物技术以及抗肿瘤化学药物的发展也必将推动单抗药物的发展与进步,单克隆抗体药物将在各种肿瘤的治疗中发挥越来越重要的作用。在未来10年内,单克隆抗体药将成为国内、外生物药品发展的主旋律。此外,利用与肿瘤细胞相关抗原的特异结合力,相应的单克隆抗体可以用于肿瘤早期诊断和预后判定。例如用放射标记抗体能够确定肿瘤存在的位置,扩散的部位和范围,以便确定手术时机和化疗方案。通过测定抗体结合白血病细胞的增减,可以检查白血病的化疗效果[sup][/sup]。利用单克隆抗体检测某些癌的特异性产物,如前列腺癌产生的酸性磷酸酶,绒毛膜上皮癌产生的促性腺激素,结肠癌产生的癌胚抗原及肝癌产生的甲胎蛋白等,有助于癌肿的早期诊断[sup][/sup]。单克隆抗体在肿瘤的治疗中的作用功不可没,但同时也面临着巨大的挑战,例如如何选择优势人群、进一步提高疗效、降低不良反应的发生都是需要进一步解决的。如贝伐单抗的突出不良反应是出血,在NCCN指南中特别指出贝伐单抗仅适用非鳞癌的[sup][/sup],既往无咯血史的患者,限制了贝伐单抗的临床应用。而其他大部分单克隆抗体均需与其他化疗药物联用,单独应用的疗效仍有限,选择合适的指标以及合适人群应用单克隆抗体仍任重而道远。[b]参考文献[/b] Adams GP, Weiner LM.Monoclonalantibody therapy of cancer .Nat Biotechnol,2005,23(9):1147~1148 甄永苏.抗肿瘤抗生素和单克隆抗体药物的研究进展.中国抗生素杂志,2002,27(1):1~5 Sievers E L, Larson R A, Stadtmauer E A, [i]et al[/i].Effica-cy and safety ofgemtuzumab ozogamicin in patients withCD33-positive acute myeloid leukemia infirst relapse .Clin Oncol,2001,19(21):3244~3246 Kamiya K, Konno H, Tanaka T, [i]et al[/i].Antitumor effect on humangastric cancer and induction of apoptosis by vascular endothelial growth factorneutralizing antibody .Jpn J Cancer Res,1999,4(21):794~798 邹学,李俊,尹庆春.单克隆抗体药物诱发肿瘤细胞凋亡的研究进展.总装备部医学学报,2008,10(2):115~117 Rao AV, Schmader K.Monoclonalantibodies as targeted therapy in hematologic malignancies in older adults .Am J GeriatrPharmacother,2007,5(3):247~250 杨海东,罗傲雪,范益军.单克隆抗体在治疗肿瘤中的研究进展.时珍国医国药,2007,18(11):2685~2686 甄红英,薛玉川,甄永苏.抗肿瘤抗生素C1027抑制血管生成及其抗肿瘤转移作用.中华医学杂志,1997,77(21):657~660 刘霆.抗肿瘤单克隆抗体靶向药物的研究进展.国外医学生理、病理科学与临床分册,2003,23(3):254~257 Plw a JL,Britta E,Jayne L,[i]et al[/i].Targeting rat anti-mouse transferrinreceptor monoclonal antibody through blood-brain barrier in mouse .pharmacology andexperiment therapeu-ties,2000,4(21):1048~1057 刘德忠,张石革.分子和抗体靶向抗肿瘤药的研究进展.中国药房,2007,18(26):2067~2068 丰雪,龙亚一,廖翰.抗肿瘤抗体-药物偶联物的临床研究进展.现代生物医学进展,2013,16(21):3164~3168 江山,杨小琼.晚期非小细胞肺癌单克隆抗体治疗的研究进展.吉林医学,2013,34(35):7482~7483 SpicerJ,Harper P.Targetedtherapies for non-small cell lung cancer .In t J C l in Pract,2005,59(9):1055~1057 彭建柳,杨丽华.人源化单克隆抗体用于肿瘤分子靶向治疗的研究进展.现代医院,2009,9(5):8~11 王飞,董军,黄强等.转基因完全人抗体的制备及其抗肿瘤作用研究.中华神经外科疾病研究杂志,2002,1(1):90~91 Kim J A.Targeted therapies for thetreatment of cancer .Am J Surg,2003,186(9):264~269 侯盛,郭亚军.单克隆抗体在肿瘤治疗中的应用.中国处方药,2007,4(61):53~56 清水惠司.抗肿瘤用药的应用及进展.临床免疫,2009,13(11):912~915 沈倍奋.抗体药物研究进展.第二军医大学学报,2002,23(10):1047~1049
[align=center]抗肿瘤单克隆抗体药物的研究进展[/align][align=center] [/align][align=center]摘 要[/align][align=center] [/align] 通过淋巴细胞杂交瘤技术或基因工程技术制备单克隆抗体药物,已经成为生物制药领域的一个重要方面,特别是对抗肿瘤单克隆抗体药物的研究已获得了重要进展。多年来,许多研究证实了抗肿瘤单克隆抗体药物的作用,为其应用于肿瘤治疗提供了重要依据。这类药物的特异性强,疗效显著。本文主要就近年来抗肿瘤单克隆抗体药物的研究进展进行了综述,并对抗肿瘤单克隆抗体药物的发展前景进行了展望。[align=left] [/align][align=left]关键词:抗肿瘤;单克隆抗体;研究进展[/align][align=center] [/align][align=center] [/align][b]一 引言[/b]抗肿瘤单抗药物因与烷化剂、抗代谢药、抗肿瘤抗生素、铂类配合物、植物药等抗肿瘤药物相比,具有高效价、高特异性、血清交叉反应少等特点与优点,在肿瘤治疗中起着不可替代的作用。单抗药物是当前生物技术药物领域甚为活跃的部分。针对特定的分子靶点(抗原),单抗有高度特异性。针对各种不同的抗原,可以制备为数众多的、各不相同的单抗;因此,作为药物来源,单抗又具有高度多样性。由于其特异性和多样性,研制单抗药物有巨大的潜力。单克隆抗体药物治疗恶性瘤主要机制有两种[sup][/sup]:一是利用单克隆抗体本身来阻断癌细胞生长的信号,单克隆抗体在癌细胞膜外与生长因子竞争结合受体,阻断信号传递过程,从而阻止癌细胞的生长和扩散,诱导细胞凋亡或者间接激活宿主的抗肿瘤免疫反应;二是利用单克隆抗体作为药物载体的靶向治疗,如将有细胞毒性的药物或有放射性的药物靶向性的运送到肿瘤细胞,从而杀伤肿瘤细胞。目前,国际上与肿瘤治疗相关的抗体研究主要集中在将抗体与耦联物作用后直接杀伤肿瘤细胞,利用抗体促进肿瘤细胞凋亡和抑制肿瘤血管生成等方面。此外,研究表明静脉内注射抗肿瘤单抗,在肿瘤部位的浓度较高,显示特异性定位;单抗与药物的偶联物通常仍保留原来单抗的分布特征,在靶肿瘤的浓度较高[sup][/sup]。[align=center]二 单克隆抗体药物作用靶点[/align]特定受体或特定的基因表达蛋白可能作为单抗药物的靶点。Rituxan是以B细胞的CD20分子作为靶点的人鼠嵌合抗体,对非霍奇金氏B细胞淋巴瘤有疗效,是第一个获美国FDA批准用于治疗恶性肿瘤的单抗。Herceptin是抗HER-2/neu癌基因编码蛋白的单抗,临床研究对乳腺癌有效,与化疗药物联合有更显著的疗效。Mylotarg是由抗CD33单抗与calicheamicin构成的偶联物,已获批准用于治疗急性复发性髓性白血病[sup][/sup]。表皮细胞生长因子受体(EGFr)在人的鳞癌、乳腺癌和脑胶质瘤等均有较高的表达。有报道,抗EGFr单抗与长春碱衍生物的偶联物在裸鼠体内试验,显示良好的抗癌效果。抗EGFr的人鼠嵌合抗体已进入临床研究。血管内皮生长因子(VEGF)在血管生成中有重要作用。据报道,抗VEGF的中和性单抗具有广谱的抗肿瘤作用,对移植于裸鼠的人体癌瘤有显著疗效[sup][/sup]。[b]三 单抗诱发肿瘤细胞凋亡[/b][align=left] 3.1 通过免疫细胞表面抗原的交联作用而诱导恶性肿瘤细胞的凋亡[/align]用于治疗血液系统恶性肿瘤的单克隆抗体药物大多是通过免疫细胞表面抗原的交联作用诱导恶性肿瘤细胞凋亡而起作用的,如目前用的抗-CD20的单克隆抗体——美罗华。其单克隆抗体的作用机制是通过诱导CD20分子在B细胞膜上的脂筏区聚集,再在一系列激酶的作用下使脂筏信号传导区域的CD20分子亲和性增强,从而形成CD20交联形式;交联的CD20分子启动了细胞内凋亡信号的传导通路,使线粒体释放出细胞色素C,激活下游的caspase级联反应,最终导致细胞凋亡[sup][/sup]。3.2 作用于恶性肿瘤细胞膜上的生长因子及其受体而诱导细胞凋亡许多生长因子及其受体通过作用于细胞的存活途径、刺激细胞的有丝分裂、促进细胞的生长增殖来阻止细胞凋亡。与正常细胞中生长因子信号传导的严格调控相比,肿瘤细胞中的失控则导致细胞的恶性增殖,从而使恶性细胞获得“永生”。单克隆抗体通过作用于恶性肿瘤细胞膜上的生长因子及其受体可阻断存活信号传导通路,从而导致其凋亡,同时还能对化疗和放疗有正协同作用。目前主要集中在对血管内皮生长因子(VEGF)及其受体、表皮生长因子受体(EGFR)等的研究。美国FDA于2006年批准了第一个用于治疗头颈部鳞状细胞癌的单克隆抗体药物——Cetuximab,它为一种IgG1单克隆抗体,主要通过干扰癌细胞表面EGFR的生长,从而减少癌细胞进入正常组织的概率,控制癌细胞的转移,达到抗癌目的[sup][/sup]。最初想到制备针对恶性肿瘤凋亡相关分子的单克隆抗体药物时,虽然从理论上来说无疑是给人们注入了一针兴奋剂,但在实际应用中则并不然,所以在通过单克隆抗体药物诱导恶性肿瘤细胞凋亡的研究和治疗中,还有待进一步开发新的、更经济、更有效地药物。[b]四 单克隆抗体耦联物[/b]4.1 抗体与化疗药物耦联目前,国内外研究较多的与单克隆抗体耦联的化学药物有平阳霉素、柔红霉素、丝裂霉素、多柔比星(阿霉素)、顺铂以及长春碱类衍生物等。同时还可以通过脂质体靶向制剂作为化疗药靶向治疗肿瘤,利用脂质体制剂将药物导向靶标进行有选择性地杀伤癌细胞和抑制癌细胞的繁殖,以达到提高疗效和高度定向作用。目前已上市的脂质体有复方氟脲嘧多相脂质体、喜树碱多相脂质体、阿霉素脂质体和紫杉醇脂质体等。4.2 抗体导向酶耦联利用抗体与肿瘤细胞表面抗原的特异性结合,将前体药物的专一性活化酶与单抗耦联,导向输入到靶细胞部分,再注入前体药物,使其在酶的作用下转化为活性药物,进而杀伤肿瘤细胞[sup][/sup]。目前这种用作前体药物的抗癌药有苦杏仁苷、氮芥、鬼臼乙叉苷、阿霉素、丝裂霉素等。而作为活化前体药物的导向酶有碱性磷酸酶、青霉素V或G酰胺酶、羧基酶肽、胸腺嘧啶核苷酶、β葡萄苷酶等。临床研究表明,单抗耦联物对于抗药性肿瘤细胞仍显示较强的杀伤活性。对由于长期使用氨甲蝶呤而出现抗药性的成骨肉瘤细胞,单抗氨甲蝶呤耦联物仍显示较强的杀伤作用。对于具有多药抗药性(MDR)的肿瘤细胞,抗P-170糖蛋白单抗构成的免疫毒素可显示选择性杀伤作用[sup][/sup]。这说明,单抗药物有可能用于克服肿瘤细胞抗药性。[b]五 单克隆抗体靶向药物[/b]单抗靶向药物是利用单抗对肿瘤表面相关抗原或特定的受体特异性识别,从而把药物直接导向肿瘤细胞,提高药物的疗效,降低药物对循环系统及其他部位的毒性。研究表明,单抗靶向药物具有很好的疗效,在免疫偶联物对移植于裸鼠的相应人体肿瘤生长有抑制作用。免疫偶联物与相应的游离物比较,具有更高更好的疗效和较低的细胞毒性[sup][/sup]。单克隆抗体体积小,能更有效地透入肿瘤;分子小、消除快、累积毒性小;所携带的弹头脱离后,可较快被清除 循环中免疫靶向结合物对靶细胞的竞争作用小;半衰期短;穿透性好;能穿过血脑屏障[sup][/sup],因而还可以作为新一代靶向载体。与化学药物、毒素、放射性核素、生物因子、基因、分化诱导剂、光敏剂、酶等物质构成单克隆抗体靶向药物,把杀伤肿瘤细胞的活性物质特异的输送到肿瘤部位,利用单抗对肿瘤表面相关抗原或特定的受体特异性识别,从而把药物直接导向肿瘤细胞,提高药物疗效,降低药物对循环系统及其他部位的毒性。近年来,随着医学、药学和生物工程学及技术的进步,临床对肿瘤的根治和对癌细胞的攻击锁定于表皮生长因子和血管内皮生长因子等靶位,使药物治疗的切入点由细胞水平提升到分子和抗体水平,从而提高了肿瘤综合治疗的效果。[align=center]六 人源化单克隆抗体[/align]单克隆抗体是近年竞相开发的品种,自1997年第1个单克隆抗体rituximab通过食品与药物管理局(FDA)批准应用于临床以来,目前已经上市的单克隆抗体靶向药物的疗效令人瞩目,在抗肿瘤、类风湿性关节炎和自身免疫系统缺陷治疗领域得到了有力的推广,其以独特的作用优势,在肿瘤的治疗中不但能够选择性杀伤癌细胞,且在体内表现出特异的分布特性,具有高效、低毒的特点,从而在生物技术产品领域中占据了1/3的市场[sup][/sup]。目前用于治疗肿瘤的单克隆抗体药已有多个,包括伊珠单抗奥加米星、帕尼单抗、曲妥珠单抗等。伊珠单抗奥加米星又名CMC-544,是以人源化抗CD22的抗体伊珠单抗与 CalichDMH偶联形成的ADC药物,用来治疗复发性或难治性B细胞非霍奇金淋巴瘤(B cell-NHL)和急性淋巴细胞白血病(ALL),目前已经进入临床 III期试验[sup][/sup]。帕尼单抗是一种IgG2单克隆抗体完全人源化可以与EGFR高度特异性地结合,进而阻断配体诱导的信号激活,从而抑制肿瘤生长。有临床研究选择既往未治疗过的ⅢB或Ⅳ期非小细胞肺癌患者比较卡铂(AUC=6,每3周),加紫杉醇(200mg/m21次/3周) 联合或不联合帕尼单抗(2.5 mg/m2,1次/周) 化疗的疗效及其安全性。研究结果显示,单纯化疗组与帕尼单抗联合化疗组之间在PFS(5.3个月对比4.2个月、P=0.55)和总生存( Overall survival,OS)(8.0个月对比8.5个月,P =0.81)上均无显著差异。结果提示帕尼单抗联合一线化疗方案可能对晚期非小细胞肺癌无明显疗效[sup][/sup]。曲妥珠单抗是一种抗Her2的单克隆抗体,他可以和肿瘤细胞的HER2/neu特异性地结合,从而阻断细胞内生长信号的转导,同时曲妥珠单抗还可以诱导体内巨噬细胞以及自然杀伤细胞攻击肿瘤细胞,以达到抑制和杀伤肿瘤细胞的目的。比较用或不用曲妥珠单抗联合一线化疗方案用以治疗ⅡB/Ⅲ期HER2/neu阳性的 NSCLC患者差异的两项大型的随机Ⅱ期临床试验,其结果显示两个试验结论相似,曲妥珠单抗不能提高化疗的疗效,但也不加重化疗的不良反应。试验中HER2/neu值为3+的患者对曲妥珠单抗治疗的反应性较好,提示曲妥珠单抗对这一较少见类型的NSCLC效果要更好[sup][/sup]。在临床治疗中使用鼠派生单抗的主要障碍之一是产生人抗鼠抗体(HAMA)反应,通过基因工程技术制备嵌合抗体的I-IPdVIA反应率较鼠源性单抗低,但完全的人源抗体才是单抗药物的发展目标。噬茵体抗体库技术和转基因小鼠技术是制备完全人源单抗的两种方法[sup][/sup]。因此,只有不断地完善单克隆抗体人源化的技术,才能更好地将完全人源化的单克隆抗体用于肿瘤分子靶向治疗中,从而使医学界迈向更高的台阶。[b]七 问题与对策[/b]在限制单克隆抗体临床治疗效果的因素有:(l)循环免疫复合物导致的肝肾功能损害。(2)可溶性肿瘤抗原释放造成的体液中的封闭作用。(3)异种蛋白反应。(4)特异性还不够专一,引起了正常细胞的伤害。(5)天然免疫功能低下(如补体介异的细胞毒,网状内皮系统清除和ADCC作用等)。(6)主要的问题还在这种免疫疗法会导致靶细胞(肿瘤细胞)上抗原的转换。为了解决这些问题,今后的研究应着重:(1)制备对肿瘤抗原有高度特异性的单克隆抗体。(2)选择不易诱导抗原转换的单克隆抗体。(3)研究副作用较少,既安全疗效又高的偶联制剂。单抗(Mab)药物存在的一个最关键问题就是人抗鼠抗体反应(HAMA)。由于用于临床研究的Mab药物一般使用鼠源Mab,这不可避免地会引起HAMA反应,所以尽量避免HAMA反应这一副作用才是Mab药物能否真正适合治疗肿瘤性疾病的重点[sup][/sup]。近些年来,Mab药物的研究主要是向减轻宿主对外源抗体的排斥,促进抗体人源化,改变抗体的氨基酸序列而增加或降低该抗体的生物学效应,加抗体的亲和力,制备双特异性抗体,改造抗体重链恒定区以增强抗体功能,以及寻找新的分子靶点(相对特异的肿瘤抗原)等方向发展[sup][/sup]。Mab药物的不断更新,必将为全球的肿瘤患者带来更大的希望。[align=center]八 总结与展望[/align]目前肿瘤治疗中使用最广泛的仍是化疗以及放射性疗法,其毒副作用较大。随着基因工程技术和DNA重组技术的兴起,利用单克隆抗体治疗肿瘤已经日渐取代副作用较大的传统疗法而成为新的发展趋向。所以,如何研制更多的单克隆抗体以及怎样更好的利用单克隆抗体治疗肿瘤,将成为肿瘤治疗研究中的又一艰巨任务。同时,生物技术以及抗肿瘤化学药物的发展也必将推动单抗药物的发展与进步,单克隆抗体药物将在各种肿瘤的治疗中发挥越来越重要的作用。在未来10年内,单克隆抗体药将成为国内、外生物药品发展的主旋律。此外,利用与肿瘤细胞相关抗原的特异结合力,相应的单克隆抗体可以用于肿瘤早期诊断和预后判定。例如用放射标记抗体能够确定肿瘤存在的位置,扩散的部位和范围,以便确定手术时机和化疗方案。通过测定抗体结合白血病细胞的增减,可以检查白血病的化疗效果[sup][/sup]。利用单克隆抗体检测某些癌的特异性产物,如前列腺癌产生的酸性磷酸酶,绒毛膜上皮癌产生的促性腺激素,结肠癌产生的癌胚抗原及肝癌产生的甲胎蛋白等,有助于癌肿的早期诊断[sup][/sup]。单克隆抗体在肿瘤的治疗中的作用功不可没,但同时也面临着巨大的挑战,例如如何选择优势人群、进一步提高疗效、降低不良反应的发生都是需要进一步解决的。如贝伐单抗的突出不良反应是出血,在NCCN指南中特别指出贝伐单抗仅适用非鳞癌的[sup][/sup],既往无咯血史的患者,限制了贝伐单抗的临床应用。而其他大部分单克隆抗体均需与其他化疗药物联用,单独应用的疗效仍有限,选择合适的指标以及合适人群应用单克隆抗体仍任重而道远。[b]参考文献[/b] Adams GP, Weiner LM.Monoclonalantibody therapy of cancer .Nat Biotechnol,2005,23(9):1147~1148 甄永苏.抗肿瘤抗生素和单克隆抗体药物的研究进展.中国抗生素杂志,2002,27(1):1~5 Sievers E L, Larson R A, Stadtmauer E A, [i]et al[/i].Effica-cy and safety ofgemtuzumab ozogamicin in patients withCD33-positive acute myeloid leukemia infirst relapse .Clin Oncol,2001,19(21):3244~3246 Kamiya K, Konno H, Tanaka T, [i]et al[/i].Antitumor effect on humangastric cancer and induction of apoptosis by vascular endothelial growth factorneutralizing antibody .Jpn J Cancer Res,1999,4(21):794~798 邹学,李俊,尹庆春.单克隆抗体药物诱发肿瘤细胞凋亡的研究进展.总装备部医学学报,2008,10(2):115~117 Rao AV, Schmader K.Monoclonalantibodies as targeted therapy in hematologic malignancies in older adults .Am J GeriatrPharmacother,2007,5(3):247~250 杨海东,罗傲雪,范益军.单克隆抗体在治疗肿瘤中的研究进展.时珍国医国药,2007,18(11):2685~2686 甄红英,薛玉川,甄永苏.抗肿瘤抗生素C1027抑制血管生成及其抗肿瘤转移作用.中华医学杂志,1997,77(21):657~660 刘霆.抗肿瘤单克隆抗体靶向药物的研究进展.国外医学生理、病理科学与临床分册,2003,23(3):254~257 Plw a JL,Britta E,Jayne L,[i]et al[/i].Targeting rat anti-mouse transferrinreceptor monoclonal antibody through blood-brain barrier in mouse .pharmacology andexperiment therapeu-ties,2000,4(21):1048~1057 刘德忠,张石革.分子和抗体靶向抗肿瘤药的研究进展.中国药房,2007,18(26):2067~2068 丰雪,龙亚一,廖翰.抗肿瘤抗体-药物偶联物的临床研究进展.现代生物医学进展,2013,16(21):3164~3168 江山,杨小琼.晚期非小细胞肺癌单克隆抗体治疗的研究进展.吉林医学,2013,34(35):7482~7483 SpicerJ,Harper P.Targetedtherapies for non-small cell lung cancer .In t J C l in Pract,2005,59(9):1055~1057 彭建柳,杨丽华.人源化单克隆抗体用于肿瘤分子靶向治疗的研究进展.现代医院,2009,9(5):8~11 王飞,董军,黄强等.转基因完全人抗体的制备及其抗肿瘤作用研究.中华神经外科疾病研究杂志,2002,1(1):90~91 Kim J A.Targeted therapies for thetreatment of cancer .Am J Surg,2003,186(9):264~269 侯盛,郭亚军.单克隆抗体在肿瘤治疗中的应用.中国处方药,2007,4(61):53~56 清水惠司.抗肿瘤用药的应用及进展.临床免疫,2009,13(11):912~915 沈倍奋.抗体药物研究进展.第二军医大学学报,2002,23(10):1047~1049
什么是诱导解离碰撞?
在原核蛋白表达体系中,如E.coli(大肠埃希菌)系统,外源基因通常需要诱导剂的诱导才能进行表达,本文详细讲述了常用诱导剂IPTG 对外源蛋白诱导表达的原理以及实验步骤。
在一定温度下,PE材料氧化诱导时间越长所代表其抗氧性越好,耐候性越好,但具体曲线或者公式是什么?为什么在PE管材涉及标准中,规定氧化诱导时间大于20min这个标准制定的依据是什么?http://simg.instrument.com.cn/bbs/images/brow/em09509.gif平时没学好,要用时候发现这个标准是根据什么制定的的不知道。各位大大帮帮忙啦!
橡胶材料有没有氧化诱导期,它和橡胶的材料的分解温度有什么关系?如何做DSC分析来确定氧化诱导期呢?
有个问题想请教大神呀,ESI源中,有没有不带电的化合物进入源里发生源内诱导解离的?解释一下呀:就是说ESI源中离子化效率比较低,源内肯定有大量的不带电的,和带电的分子,当电压大,进行源内诱导解离时,带电的能被打碎这点没有问题,我的问题是不带电的化合物也能被打碎吗?如果这样的话,化合物发生异裂,不就产生两个离子,与加氢带正电的准分子离子同一位置均裂的话,碎片分子量不就差一了吗?这其中还涉及到一些其他的问题,谢谢各位大神呀!还有,就是谁有关于讲ESI源质谱解析的书推荐的呀?《电喷雾质谱应用技术》就算了,那本书主要侧重行业应用,讲的范围比较广,我想知道的是关于解析这块的,现在好多书都是以EI为基础的讲解,感觉与平时实验有点不一样,谢谢!
常吃这7类食物易患肿瘤第一种:过食温热性食物。 所谓的温热性食物,一是指直接感受到的温度过高、过烫的食品;一是指属于中医药性划分中的温热之性食物。因为过于辛热,容易损伤消化道黏膜上皮细胞,所以这类性质的食品最易诱发的是消化系统的肿瘤。过烫饮食是导致食道癌等消化道肿瘤发生的重要原因。饮食过热,会损伤、刺激食道黏膜上皮,长期刺激下将诱导组织恶变。第二种:酒。 酒为辛热之性,长期饮酒,是消化系统肿瘤发病最常见的因素。山东西南地区为食道癌的高发区,就与这一地区饮食中嗜热、嗜酒有关。其实,早在1964年,世界卫生组织就认为酒精饮料的过度消费与口腔癌、喉癌及食道癌有关。在分别对江苏启东县肝癌相对高发的通兴乡与相对低发的西宁乡进行的调查中发现:前者肝癌病死率是后者的2.5倍。而两乡只有一路之隔,自然环境基本相同,吸烟率也相当,所不同的是肝癌高发区的人们爱喝酒,低发区的人们则喜欢饮茶。第三种:煎炸、熏烤的食物。 同样属于中医热性食物的还有煎炸、熏烤的食物,这类食物因经过高温的油炸和烤制,亦随之变为热性食品。经研究证实,长期食用此类食品,易于导致胃癌、肠癌、肝癌等。第四种:咖啡。 就像中国人嗜茶一样,西方人多喜喝咖啡。美国学者在经过5年的流行病学调查后认为,咖啡因对胰腺癌和膀胱癌的形成有一定影响。经常饮咖啡的人比不饮咖啡的人患这两种癌的可能性大2~3倍。在胰腺癌患者中,至少一半是由于过多饮用咖啡引起的。第五种:咸味太过的食物。 除了食盐,咸味食物还包括咸菜、咸鱼、咸肉以及其他腌制食品等。根据相关研究显示,常食腌制食品者食管癌发病的危险性是不常食用者的2.79倍。嗜食咸味食物最容易导致的癌症是胃癌。嗜食咸鱼的日本渔民,胃癌、食道癌的发生较为普遍。嗜食咸鱼亦是致鼻咽癌的一个重要因素。马来西亚研究人员发现,沿海居民患鼻咽癌较多,原因与他们爱吃咸鱼有关。我国普查资料证明,在胃癌高发区,人均每天摄人食盐50克;而胃癌低发区,人均食盐摄入量仅为6克左右。 第六种:霉烂食物。 黄曲霉毒素已被公认为是最强烈的致癌物,而黄曲霉菌富含于霉烂的谷物、玉米、花生中,该菌在温暖、潮湿的环境下易于生长繁殖,研究发现其可诱发实验动物的多种肿瘤。这种因素是导致亚洲、非洲某些地区癌肿高发的重要原因。第七种:腌制食品。 亚硝酸盐也是公认的致癌物,蔬菜、鲜肉等腌制后会产生大量的亚硝酸盐。盐腌的干鱼中,发酵的腌菜、泡菜、酸菜中,隔夜的煮熟白菜、香肠、肉类中都含有亚硝基化合物,如果经常食用这些食物就有致癌的危险。
[size=15px][b][font=&][color=#0070c0]1[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]GA[/color][/font][font=宋体][color=#0070c0]诱导[/color][/font][font=&][color=#0070c0]KRAS[/color][/font][font=宋体][color=#0070c0]降解[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]作者首先通过筛选了一个自制的化合物库,用含有[/font][font=&] KRAS[sup]G12A[/sup][/font][font=宋体]突变的[/font][font=&]MM[/font][font=宋体]细胞系[/font][font=&]MM.1S[/font][font=宋体]和[/font][font=&]RPMI 8226[/font][font=宋体]鉴定可以降低[/font][font=&]KRAS[/font][font=宋体]水平的化合物,发现藤黄酸([/font][font=&]Gambogic acid[/font][font=宋体],[/font][font=&]GA[/font][font=宋体],)处理可降低两种细胞系中[/font][font=&]KRAS[/font][font=宋体]的水平,而其他化合物没有表现出相似或较弱的效果。进一步实验表明,[/font][font=&]GA[/font][font=宋体]可以以浓度和时间依赖性方式降低两种细胞系中[/font][font=&]KRAS[/font][font=宋体]的水平,以及[/font][font=&] KRAS[/font][font=宋体]的下游[/font][font=&]p-ERK[/font][font=宋体]([/font][font=&]MAPK[/font][font=宋体]通路)和[/font][font=&]p-AKT[/font][font=宋体]([/font][font=&]PI3K[/font][font=宋体]通路)。此外,[/font][font=&]q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url][/font][font=宋体]显示[/font][font=&]GA[/font][font=宋体]不会改变[/font][font=&]KRAS[/font][font=宋体]的转录水平,表明[/font][font=&]GA[/font][font=宋体]可能在转录后水平降低[/font][font=&]KRAS[/font][font=宋体]。[/font][font=&][/font][/size][size=15px][font=宋体]进一步使用蛋白酶体抑制剂[/font][font=&]MG132[/font][font=宋体]发现[/font][font=&]MG132 [/font][font=宋体]部分挽救了[/font][font=&]GA[/font][font=宋体]诱导的[/font][font=&]KRAS[/font][font=宋体]蛋白下调,表明[/font][font=&] KRAS [/font][font=宋体]蛋白降解与泛素[/font][font=&]-[/font][font=宋体]蛋白酶体途径有关,此外,我们采用放线菌酮([/font][font=&]CHX[/font][font=宋体])测定评估显示[/font][font=&]GA[/font][font=宋体]处理后,[/font][font=&]KRAS[/font][font=宋体]的半衰期显著缩短。这些数据表明[/font][font=&]GA[/font][font=宋体]可以诱导[/font][font=&]KRAS[/font][font=宋体]的蛋白酶体降解,进而损害[/font][font=&]MM[/font][font=宋体]细胞中的下游信号转导[/font][font=宋体] [size=15px][b]2、鉴定USP2作为GA的新靶标[/b][/size][size=15px]为了确定GA诱导KRAS降解的靶标,作者合成了生物素标记的GA,发现它保留了在MM细胞中诱导KRAS降解的能力,进一步利用该探针开展Pulldown+MS,鉴定到的互作蛋白中,HSP90和USP2这两种与蛋白质降解相关。HSP90是一个已知的GA靶点,作为伴侣蛋白,它可以调节各种蛋白的稳定性,然而HSP90抑制剂STA9090处理不能降低MM.1S细胞中KRAS的蛋白水平,表明KRAS不是 HSP90的底物。因此,作者更加关注USP2,一种新型的GA互作蛋白。通过Pulldown+WB、免疫荧光共定位、竞争实验、CETSA、DARTS实验共同证实了USP2是GA的直接相互作用蛋白。此外,GA在体外相对特异性地抑制USP2的去泛素化活性( [/size][size=15px][b]3、半胱氨酸284对于GA与USP2的共价结合至关重要[/b][/size][size=15px]然后,作者研究了GA和USP2之间的相互作用模式,发现碘乙酸(IAA,一种半胱氨酸烷化剂)与USP2的预孵育完全抑制了GA与USP2的结合,表明GA可能与USP2的半胱氨酸共价结合,这与之前的报道一致,即GA可以通过半胱氨酸残基与其靶标共价结合。接着,通过USP2蛋白与GA一起孵育后质谱鉴定,发现Cys284残基被GA共价修饰。此外,通过构建USP2的两个突变体(C276S、C284S),发现GA 与 USP2(C276S)结合,而不是USP2(C284S),表明GA可以与USP2的Cys284残基特异性形成共价键。结合动力学显示GA 以剂量和时间依赖性方式与 USP2 共价结合,分子对接显示P565和 R289对于形成GA与USP2结合的口袋至关重要。同样通过蛋白点突变实验发现P565 和 R289对GA-USP2相互作用至关重要 [/size][size=15px][b]4、USP2调节KRAS的稳定性[/b][/size][size=15px]前面发现GA降低MM细胞中KRAS蛋白水平并抑制USP2活性,表明USP2可能调节KRAS的稳定性。作者利用CRISPR/Cas9敲除MM.1S和RPMI 8226细胞系中的USP2,发现敲除USP2导致KRAS蛋白水平降低,mRNA水平不变, KRAS的下游效应子p-ERK水平也显著降低。此外,敲除诱导的KRAS下调可被两种细胞系中的蛋白酶体抑制剂MG132挽救。结果表明USP2可以增强KRAS的蛋白质稳定性。进一步研究发现USP2与KRAS互作并使KRAS去泛素化,表明KRAS是USP2的底物 [/size][size=15px][b]5、USP2敲除抑制MM细胞的增殖[/b][/size][size=15px]鉴于KRAS在MM细胞增殖和存活中的关键作用,作者假设USP2敲除可能会使KRAS不稳定,从而导致MM细胞增殖抑制。结果显示USP2敲除显著抑制MM.1S和RPMI 8226细胞系的增殖,诱导这两种MM细胞系凋亡。此外,MM.1S 细胞的异种移植 MM 模型发现,USP2敲除显著抑制肿瘤生长,免疫组化染色也显示USP2敲除组的KRAS水平降低,这些数据共同表明USP2在MM细胞的增殖和存活中起着关键作用 [/size][size=15px][b]6、GA通过靶向USP2和破坏KRAS的稳定性来诱导MM细胞凋亡[/b][/size][size=15px]据报道,GA通过抑制PI3K/Akt/mTOR、[i]NF-κ[/i]B 和其他信号通路诱导MM细胞凋亡。作者发现GA处理可以以剂量依赖性方式降低MM.1S和RPMI 8226细胞的活力并促进细胞凋亡。此外,过表达USP2的细胞对GA诱导的活力降低和KRAS降解表现出部分抗性,且GA处理可以提高 KRAS的泛素化水平。KRAS[sup]G12C[/sup]的过表达可以部分消除USP2敲除诱导的细胞生长抑制。这些数据表明,GA在 MM 细胞中诱导的细胞毒性至少部分归因于其靶向USP2,这降低了KRAS的稳定性。[/size][size=15px]最后,作者探讨了USP2在MM中的临床意义。通过GSE13591数据集发现MM患者骨髓样本中的USP2 mRNA水平明显更高。此外,作者发现原发性骨髓瘤细胞和骨髓瘤细胞系中USP2的蛋白水平高于正常外周血单核细胞(PBMC)和正常骨髓活检,且高USP2水平的MM患者的总生存期降低,这些结果共同表明USP2表达增加与MM中较差的结局之间存在很强的相关性,表明USP2可能是MM治疗的有前途的靶点。[/size][/font][font=&][/font][/size]
大家好!小弟想用TA的Q100 DSC测定物质的氧化诱导时间,请问我该怎么弄?最好能给小弟举个例子!!小弟先谢过了,呵呵
请问,有做LIBS(激光诱导击穿光谱)或者LIPS(激光诱导等离子光谱)研究的朋友吗
氧化诱导时间测试时需要发生化学变化放气等,做此测试是否对DSC仪器污染很大?
请教大家,原理上是不是如果被测物没有荧光,只要抬高荧光背景,电泳时就会出负峰呢?我测的4个不同的物质,都没有荧光,在同一个体系跑CZE-间接激光诱导荧光,发现出的色谱图相同,不知怎么破解?求大神科普一下间接激光诱导荧光的知识和经验
如题,哪位使用过安捷伦6120的“碰撞诱导电离”功能?对于里面的“质量”/“碰撞诱导解离电压对”的参数设置有没有什么特别的讲究的?
最近在学习用DSC作氧化诱导期,仪器是TA Q200,我想问一下随机带热分析软件可以自动分析氧化诱导期吗?还是在实验进行时选择自动分析选项就可以了?新手,很多东西都不懂,还望大家多多帮忙![em06]







 我要推广仪器
我要推广仪器
 下载APP
下载APP