拉力试验机又称万能材料试验机。可以在受控的速度下对样条进行伸展,弯曲,压缩,甚至穿刺,直到使他们断裂。 万能材料试验机越来越常出现在塑料注塑和挤出业的实验室中,它们越来越多地运用到前沿产品的工艺开发。还有一个原因是它们对进料和成品的质量控制有着更为严格的监控。美国chatillon的产品是一个很好的实例。 许多OEM厂家,特别是那些医疗装置或汽车领域的,需要塑料加工商在生产运转结束时自行进行测试。内部测试的另一个原因是改善工艺控制,这能降低废品率和实现真正的回报。 电子拉力机的规格由框架能够承受的最大负载和承载单元的最大负载结合起来进行表示。负载单元安装在电机驱动或油压驱动的移动横梁上。与夹具相连的承载单元测量力,可以从数字显示或电脑上读数。许多电子拉力机具有可互换的传感器,从而能与待测试材料匹配。
据国外媒体报道,一个工程师组研发出一种新型弹性电池,它将为大量医学与技术的进步铺平道路。 它由众多可弯曲和延伸到最大程度的小电池构成,可令医生直接将其缝合到心脏组织中。一旦全部完成,这种新电池还将导致技术改进。这种弹性电池的研发人之一、美国西北大学的黄永刚(Yonggang Huang音译)表示:“这样的可伸缩电池使弹性电子集成在一个小包中。”这种小电池可从正常结构扭曲最大300%,而且不会遭到破坏。 《自然通讯》杂志报道,用于这类电池的无线电力传输系统将使这些电池无需直接接触就可充电。黄永刚说:“伸展这种电池时,波状连接线就会散开,但存储部件几乎保持原状,不会变形,因为它们比连接线有更大硬度。” 从理论上说,这种电池将来还能用于目前正被试验的弹性手机。这项新技术由美国西北大学的黄永刚(左)和伊利诺斯大学的约翰-罗杰斯等工程师研发。罗杰斯以前凭借这项特殊技术荣获麻省理工学院莱梅尔森奖,获得50万美元奖金。它在弹性技术领域并非第一个创新产品。2010年研发成功的LED灯板已应用在医学领域。
红外光谱(infrared spectra),以波长或波数为横坐标以强度或其他随波长变化的性质为纵坐标所得到的反映红外射线与物质相互作用的谱图。按红外射线的波长范围,可粗略地分为[url=https://insevent.instrument.com.cn/t/1p][color=#3333ff]近红外光谱[/color][/url](波段为0.8~2.5微米)、中红外光谱(2.5~25微米)和远红外光谱(25~1000微米)。对物质自发发射或受激发射的红外射线进行分光,可得到红外发射光谱,物质的红外发射光谱主要决定于物质的温度和化学组成;对被物质所吸收的红外射线进行分光,可得到红外吸收光谱。每种分子都有由其组成和结构决定的独有的红外吸收光谱,它是一种分子光谱。分子的红外吸收光谱属于带状光谱。原子也有红外发射和吸收光谱,但都是线状光谱。量子场论或量子电动力学可以正确地描述和解释红外射线(一种电磁辐射)与物质的相互作用。若采用半经典的理论处理方法,即对组成物质的分子和原子作为量子力学体系来处理,辐射场作为一种经典物理中的电磁波并忽略其光子的特征,则分子红外光谱是由分子不停地作振动和转动而产生的。分子振动是指分子中各原子在平衡位置附近作相对运动,多原子分子可组成多种振动模式。当孤立分子中各原子以同一频率、同一相位在平衡位置附近作简谐振动时,这种振动方式称简正振动。含N个原子的分子应有3N-6个简正振动方式;如果是线性分子,只有3N-5个简正振动方式。图中示出非线性3原子分子仅有的3种简正振动模式。分子的转动指的是分子绕质心进行的运动。分子振动和转动的能量不是连续的,而是量子化的。当分子由一种振动(或转动)状态跃迁至另一种振动(或转动)状态时,就要吸收或发射与其能级差相应的光。研究红外光谱的方法主要是吸收光谱法。使用的光谱有两种类型。一种是单通道或多通道测量的棱镜或光栅色散型光谱仪,另一种是利用双光束干涉原理并进行干涉图的傅里叶变换数学处理的非色散型的傅里叶变换红外光谱仪。红外光谱具有高度的特征性,不但可以用来研究分子的结构和化学键,如力常数的测定等,而且广泛地用于表征和鉴别各种化学物种。 红外识谱歌红外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
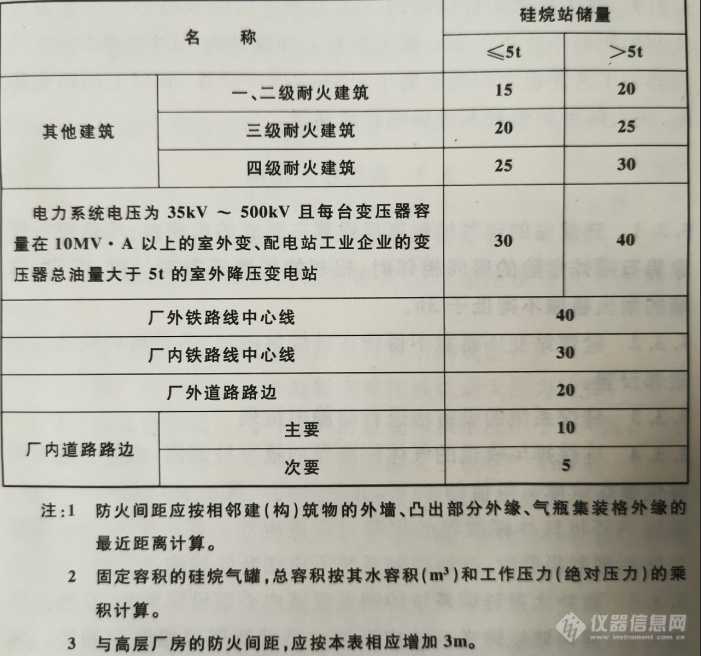
[b]硅烷站土建结构布置探讨[/b]硅烷站,是指放置硅烷或硅烷混合气体的钢瓶、钢瓶集装格、Y型钢瓶、长管拖车或ISO标准集装瓶组、硅烷气化装置、尾气处理装置、电气装置等,并通过管道向生产厂房供应硅烷气体的独立建筑物或区域。硅烷由于其极活泼的化学性质,在-180℃都会和氧气发生剧烈的化学反应,被认为是最具危险性质的特种气体。硅烷站作为硅烷的集中放置点,危险性不言而喻。[img=,690,645]https://ng1.17img.cn/bbsfiles/images/2019/09/201909101317331870_6615_3989203_3.png!w690x645.jpg[/img][img=,690,645]https://ng1.17img.cn/bbsfiles/images/2019/09/201909101317331870_6615_3989203_3.png!w690x645.jpg[/img][img=,690,645]https://ng1.17img.cn/bbsfiles/images/2019/09/201909101317331870_6615_3989203_3.png!w690x645.jpg[/img][b]建筑结构布置:[/b]硅烷站应该布置在工厂常年最小频率风向的下风侧,并应远离有明火或散发火花的地点,不得布置在人员密集地段或主要交通要到的临近处。硅烷站应采用单层钢筋混凝土或钢框架、排架结构,钢框架、排架结构应采用防火保护措施。硅烷站应设置不燃烧体的实体围墙,其高度不应小于2.5m。大宗硅烷系统设备必须布置在独立的开敞式建筑或空旷区域,不得建在地下室。当采用开敞式建筑结构形式时,硅烷站立柱和墙面遮挡部分面积不得大于建筑外围面积的25%。四周有障碍物时,硅烷站与障碍物的距离应大于障碍物高度的2倍。[b]结语:[/b]硅烷站的设置应方便运输车辆和消防车辆的进出,硅烷站的储存、分配区域应设有防止车辆撞击的保护措施。总而言之,硅烷站的设计、施工都要非常规范、谨慎,考虑到结合环境的一切可能因素,才能安全化生产。
结构分子生物的发展梁栋材 (中科院生物物理所,生物大分子国家实验室,北京,100101) 伦琴发现X射线后的一百年间,X射线在物质结构研究上立下了永不磨灭的伟大功绩。1912年劳埃发现晶体的X射线衍射,开创了晶态物质结构的新纪元。仅隔了22年Bernal和Crowfoot在1934年就成功地拍摄到第一张蛋白质 (胃蛋白酶) 单晶体的X射线衍射照片。事隔21年后的1953年Perutz发现了同晶置换法可以解决生物大分子晶体结构测定中的相位问题,从而蛋白质晶体学开始踏上自己发展的伟大历程。在1957年和1959年Kendrew和Perutz分别获得了肌红蛋白和血红蛋白的低分辨率 (6?和5?) 结构,在此期间Watson和Criek共同建立了DNA双螺旋的结构模型。他们的伟大成就为分子生物学奠定了基础。从1957年到1967年的十年里,随着溶菌酶结构之后,胰凝乳蛋白酶A、核糖核酸酶、核糖核酸酶S和羧肽酶也分别获得了高分辨率的结果,表明蛋白质晶体学已经成为一门成熟的学科。从六十年代末进入七十年代,蛋白质晶体学从对生物大分子三维结构测定迈入生物大分子三维结构与其生物学功能之间的关系研究,从而它既是分子生物学研究的有力的重要手段,同时也开始为结构分子生物学的建立和发展历程创造着条件。生物大分子发挥其生物学功能必需具备:(1) 稳定的特征的三维结构,(2) 其三维结构在各个水平上的运动。 随着学科的交叉渗透和迅速发展,一个极其重要的分支学科--结构分子生物学正在高速发展,并已成为当前生物学中的一个重要前沿学科。结构分子生物学是结构生物学中的一个最重要、最活跃的研究层次,它是在分子层次上从结构角度特别是从三维结构的角度研究和阐明当前生物学中各个前沿领域的重要学科问题。结构分子生物学是一个包括生物、物理、化学和计算数学等多学科交叉的前沿,其中心任务就是生物大分子的结构与其生物功能关系的研究。 结构分子生物学对生物大分子 (包括多亚基、多分子的复合物及复杂的复合体) 三维结构及其运动的研究手段主要有:X射线晶体衍射--蛋白质晶体学,二维及多维核磁共振谱,电子晶体学及电镜三维重组,中子衍射,其他包括应用傅里叶变换技术的各种谱学方法。他们都各有自身特有的优越性和不足。然而,无论从已测定生物大分子三维结构的数量上、精确度上或其发展潜力上,X射线单晶衍射方法--蛋白质晶体学--至今及可见的将来仍将占统治地位,都是其他手段不可相比的和不可代替的生物大分子三维结构研究手段。八十年代迅速崛起的蛋白质工程及药物设计已充分显示蛋白质晶体学方法是处于不可取代的重要地位,也表明结构分子生物学是生物高技术应用研究的重要前提和保证。 在结构分子生物学领域中由于学科上的重大突破和杰出成变而荣获诺贝尔奖金的科学家,前后有:F. H. C. Crick和J. D. Watson (1962年生理与医学奖),M. F. Perutz和J. C. Kendrew (1962年化学奖),D. C. Hodgkin (1964年化学奖),A. Klug (1982年化学奖)和R. Huber (1988年化学奖) 等,他们都是蛋白质晶体学家。 当前结构分子生物学在国际上的发展趋势有如下几个方面的特点: 以蛋白质为主要手段的生物大分子三维结构测定在高速度发展。 结构分子生物学的迅速兴起和发展,它在近些年来的生物物理学研究中已经毫无疑问地占据了主流的位。 结构分子生物学的研究成果越来越受到生命科学各个领域的重视和引用。 国际上很多难度高、意义重大的三维结构均是以蛋白质晶体学手段在近几年突破的。 结构研究已由单一分子进入研究分子之间相互作用的复合物和分子体系的结构。 蛋白质晶体学研究从生物大分子静态 (时间统计) 的结构分析开始进入了动态 (时间分辨) 的结构研究及力学分析。 技术和方法高速发展。 基础研究不断深入与扩展的同时,应用研究在迅速发展。 激烈的竞争机制已打破了传统的学院工的研究体制和格局。
红外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。 样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。 2960、2870是甲基,2930、2850亚甲峰。 1470碳氢弯,1380甲基显。 二个甲基同一碳,1380分二半。 面内摇摆720,长链亚甲亦可辨。 烯氢伸展过三千,排除倍频和卤烷。 末端烯烃此峰强,只有一氢不明显。 化合物,又键偏,~1650会出现。 烯氢面外易变形,1000以下有强峰。 910端基氢,再有一氢990。 顺式二氢690,反式移至970; 单氢出峰820,干扰顺式难确定。 炔氢伸展三千三,峰强很大峰形尖。 三键伸展二千二,炔氢摇摆六百八。 芳烃呼吸很特征,1600~1430。 1650~2000,取代方式区分明。 900~650,面外弯曲定芳氢。 五氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢 醇酚羟基易缔合,三千三处有强峰。 C-O伸展吸收大,伯仲叔醇位不同。 1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。 1110醚链伸,注意排除酯酸醇。 若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。 苯环若有甲氧基,碳氢伸展2820。 次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。 缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。 羰基伸展一千七,2720定醛基。 吸电效应波数高,共轭则向低频移。 张力促使振动快,环外双键可类比。 二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。 羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。 1740酯羰基,何酸可看碳氧展。 1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。 1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。 羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。 伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。
红外识谱歌外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
椭偏仪结构原理与发展[img]http://www.instrument.com.cn/bbs/images/affix.gif[/img][url=http://www.instrument.com.cn/bbs/download.asp?ID=63165]椭偏仪结构原理与发展[/url]
电子引伸计结构以及工作原理金属材料试验机电子引伸计是感受试件变形的传感器,应变计式的引伸计由于原理简单、安装方便,目前是广泛使用的一种类型。引伸计按测量对象,可分为轴向引伸计、横向引伸计、夹式引伸计。钢筋拉力机电子引伸计径向引伸计:用于检测标准试件径向收缩变形,它与轴向引伸计配合用来测定泊松比μ,它将径向变形(或横向某一方向的变形)变换成电量,再通过二次仪表测量、记录或控制另一设备。铝型材拉力试验机电子引伸计夹式引伸计 用于检测裂纹张开位移。电子引伸计是断裂力学实验中最常用的仪器之一,它较多用在测定材料断裂韧性实验中。精度高,安装方便、操作简单。试件断裂时引伸计能自动脱离试件,适合静、动变形测量。金属材料弹性模量试验机电子引伸计应变片、变形传递杆、弹性元件、限位标距杆、刀刃和夹紧弹簧等。测量变形时, 将引伸计装卡于试件上, 刀刃与试件接触而感受两刀刃间距内的伸长,通过变形杆使弹性元件产生应变, 应变片将其转换为电阻变化量, 再用适当的测量放大电路转换为电压信号。电子万能材料试验机电子引伸计一般是用于,金属材料试验机,金属拉力试验机,万能材料试验机,伺服材料试验机,铝型材材料试验机,钢材拉力试验机,钢筋拉力机等等
红外光谱的应用速记红外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。 样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。 2960、2870是甲基,2930、2850亚甲峰。 1470碳氢弯,1380甲基显。 二个甲基同一碳,1380分二半。 面内摇摆720,长链亚甲亦可辨。 烯氢伸展过三千,排除倍频和卤烷。 末端烯烃此峰强,只有一氢不明显。 化合物,又键偏,~1650会出现。 烯氢面外易变形,1000以下有强峰。 910端基氢,再有一氢990。 顺式二氢690,反式移至970; 单氢出峰820,干扰顺式难确定。 炔氢伸展三千三,峰强很大峰形尖。 三键伸展二千二,炔氢摇摆六百八。 芳烃呼吸很特征,1600~1430。 1650~2000,取代方式区分明。 900~650,面外弯曲定芳氢。 五氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢 醇酚羟基易缔合,三千三处有强峰。 C-O伸展吸收大,伯仲叔醇位不同。 1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。 1110醚链伸,注意排除酯酸醇。 若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。 苯环若有甲氧基,碳氢伸展2820。 次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。 缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。 羰基伸展一千七,2720定醛基。 吸电效应波数高,共轭则向低频移。 张力促使振动快,环外双键可类比。 二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。 羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。 1740酯羰基,何酸可看碳氧展。 1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。 1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。 羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。 伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。 胺尖常有干扰见,N-H伸展三千三, 叔胺无峰仲胺单,伯胺双峰小而尖。 1600碳氢弯,芳香仲胺千五偏。 八百左右面内摇,确定最好变成盐。 伸展弯曲互靠近,伯胺盐三千强峰宽, 仲胺盐、叔胺盐,2700上下可分辨, 亚胺盐,更可怜,2000左右才可见。 硝基伸缩吸收大,相连基团可弄清。 1350、1500,分为对称反对称。 氨基酸,成内盐,3100~2100峰形宽。 1600、1400酸根展,1630、1510碳氢弯。 盐酸盐,羧基显,钠盐蛋白三千三。 矿物组成杂而乱,振动光谱远红端。 钝盐类,较简单,吸收峰,少而宽。 注意羟基水和铵,先记几种普通盐。 1100是硫酸根,1380硝酸盐, 1450碳酸根,一千左右看磷酸。 硅酸盐,一峰宽,1000真壮观。 勤学苦练多实践,红外识谱不算难。
外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。 样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。 2960、2870是甲基,2930、2850亚甲峰。 1470碳氢弯,1380甲基显。 二个甲基同一碳,1380分二半。 面内摇摆720,长链亚甲亦可辨。 烯氢伸展过三千,排除倍频和卤烷。 末端烯烃此峰强,只有一氢不明显。 化合物,又键偏,~1650会出现。 烯氢面外易变形,1000以下有强峰。 910端基氢,再有一氢990。 顺式二氢690,反式移至970; 单氢出峰820,干扰顺式难确定。 炔氢伸展三千三,峰强很大峰形尖。 三键伸展二千二,炔氢摇摆六百八。 芳烃呼吸很特征,1600~1430。 1650~2000,取代方式区分明。 900~650,面外弯曲定芳氢。 五氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢 醇酚羟基易缔合,三千三处有强峰。 C-O伸展吸收大,伯仲叔醇位不同。 1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。 1110醚链伸,注意排除酯酸醇。 若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。 苯环若有甲氧基,碳氢伸展2820。 次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。 缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。 羰基伸展一千七,2720定醛基。 吸电效应波数高,共轭则向低频移。 张力促使振动快,环外双键可类比。 二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。 羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。 1740酯羰基,何酸可看碳氧展。 1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。 1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。 羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。 伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。 胺尖常有干扰见,N-H伸展三千三, 叔胺无峰仲胺单,伯胺双峰小而尖。 1600碳氢弯,芳香仲胺千五偏。 八百左右面内摇,确定最好变成盐。 伸展弯曲互靠近,伯胺盐三千强峰宽, 仲胺盐、叔胺盐,2700上下可分辨, 亚胺盐,更可怜,2000左右才可见。 硝基伸缩吸收大,相连基团可弄清。 1350、1500,分为对称反对称。 氨基酸,成内盐,3100~2100峰形宽。 1600、1400酸根展,1630、1510碳氢弯。 盐酸盐,羧基显,钠盐蛋白三千三。 矿物组成杂而乱,振动光谱远红端。 钝盐类,较简单,吸收峰,少而宽。 注意羟基水和铵,先记几种普通盐。 1100是硫酸根,1380硝酸盐, 1450碳酸根,一千左右看磷酸。 硅酸盐,一峰宽,1000真壮观。 勤学苦练多实践,红外识谱不算难。
上网发现了这个资料不错,转载过来和大家分享....过几天在转到红外版去吧,我们先学习[em0801][em0801][em0801][color=#DC143C][size=4]红外识谱歌[/size][/color][color=#00008B]外可分远中近,中红特征指纹区1300来分界,注意横轴划分异看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系识图先学饱和烃,三千以下看峰形2960、2870是甲基,2930、2850亚甲峰1470碳氢弯,1380甲基显二个甲基同一碳,1380分二半面内摇摆720,长链亚甲亦可辨烯氢伸展过三千,排除倍频和卤烷末端烯烃此峰强,只有一氢不明显化合物,又键偏,~1650会出现 烯氢面外易变形,1000以下有强峰910端基氢,再有一氢990顺式二氢690,反式移至970单氢出峰820,干扰顺式难确定炔氢伸展三千三,峰强很大峰形尖. 三键伸展二千二,炔氢摇摆六百八芳烃呼吸很特征,1600~1430 1650~2000,取代方式区分明"900~650,面外弯曲定芳氢五氢吸收有两峰,700和750四氢只有750,二氢相邻830间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰, C-O伸展吸收大,伯仲叔醇位不同1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚1110醚链伸,注意排除酯酸醇若与'pai"键紧相连,二个吸收要看准. 1050对称峰,1250反对称苯环若有甲氧基,碳氢伸展2820次甲基二氧连苯环,930处有强峰环氧乙烷有三峰,1260环振动九百上下反对称,八百左右最特征缩醛酮,特殊醚,1110非缩酮酸酐也有C-O键,开链环酐有区别开链强宽一千一,环酐移至12500 羰基伸展一千七,2720定醛基吸电效应波数高,共轭则向低频移张力促使振动快,环外双键可类比二千五到三千三,羧酸氢键峰形宽920,钝峰显,羧基可定二聚酸酸酐千八来偶合,双峰60严相隔链状酸酐高频强,环状酸酐高频弱羧酸盐,偶合生,羰基伸缩出双峰 1600反对称,1400对称峰1740酯羰基,何酸可看碳氧展1180甲酸酯,1190是丙酸1220乙酸酯,1250芳香酸1600兔耳峰,常为邻苯二甲酸氮氢伸展三千四,每氢一峰很分明羰基伸展酰胺I,1660有强峰N-H变形酰胺II,1600分伯仲伯胺频高易重叠,仲酰固态15505 碳氮伸展酰胺III,1400强峰显胺尖常有干扰见,N-H伸展三千三叔胺无峰仲胺单,伯胺双峰小而尖1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐伸展弯曲互靠近,伯胺盐三千强峰宽仲胺盐、叔胺盐,2700上下可分辨亚胺盐,更可怜,2000左右才可见硝基伸缩吸收大,相连基团可弄清1350、1500,分为对称反对称 氨基酸,成内盐,3100~2100峰形宽1600、1400酸根展,1630、1510碳氢弯盐酸盐,羧基显,钠盐蛋白三千三.矿物组成杂而乱,振动光谱远红端 钝盐类,较简单,吸收峰,少而宽注意羟基水和铵,先记几种普通盐1100是硫酸根,1380硝酸盐1450碳酸根,一千左右看磷酸硅酸盐,一峰宽,1000真壮观勤学苦练多实践,红外识谱不算难[/color]
外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。 样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。 2960、2870是甲基,2930、2850亚甲峰。 1470碳氢弯,1380甲基显。 二个甲基同一碳,1380分二半。 面内摇摆720,长链亚甲亦可辨。 烯氢伸展过三千,排除倍频和卤烷。 末端烯烃此峰强,只有一氢不明显。 化合物,又键偏,~1650会出现。 烯氢面外易变形,1000以下有强峰。 910端基氢,再有一氢990。 顺式二氢690,反式移至970; 单氢出峰820,干扰顺式难确定。 炔氢伸展三千三,峰强很大峰形尖。 三键伸展二千二,炔氢摇摆六百八。 芳烃呼吸很特征,1600~1430。 1650~2000,取代方式区分明。 900~650,面外弯曲定芳氢。 五氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢。 醇酚羟基易缔合,三千三处有强峰。 C-O伸展吸收大,伯仲叔醇位不同。 1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。 1110醚链伸,注意排除酯酸醇。 若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。 苯环若有甲氧基,碳氢伸展2820。 次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。 缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。 羰基伸展一千七,2720定醛基。 吸电效应波数高,共轭则向低频移。 张力促使振动快,环外双键可类比。 二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。 羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。 1740酯羰基,何酸可看碳氧展。 1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。 1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。 羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。 伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。 胺尖常有干扰见,N-H伸展三千三, 叔胺无峰仲胺单,伯胺双峰小而尖。 1600碳氢弯,芳香仲胺千五偏。 八百左右面内摇,确定最好变成盐。 伸展弯曲互靠近,伯胺盐三千强峰宽, 仲胺盐、叔胺盐,2700上下可分辨, 亚胺盐,更可怜,2000左右才可见。 硝基伸缩吸收大,相连基团可弄清。 1350、1500,分为对称反对称。 氨基酸,成内盐,3100~2100峰形宽。 1600、1400酸根展,1630、1510碳氢弯。 盐酸盐,羧基显,钠盐蛋白三千三。 矿物组成杂而乱,振动光谱远红端。 钝盐类,较简单,吸收峰,少而宽。 注意羟基水和铵,先记几种普通盐。 1100是硫酸根,1380硝酸盐, 1450碳酸根,一千左右看磷酸。 硅酸盐,一峰宽,1000真壮观。 勤学苦练多实践,红外识谱不算难。
红外识谱歌外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。[em17] [em17]
[转载] 红外识谱歌外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
红外识谱歌红 外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。 样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。 2960、2870是甲基,2930、2850亚甲峰。 1470碳氢弯,1380甲基显。 二个甲基同一碳,1380分二半。 面内摇摆720,长链亚甲亦可辨。 烯氢伸展过三千,排除倍频和卤烷。 末端烯烃此峰强,只有一氢不明显。 化合物,又键偏,~1650会出现。 烯氢面外易变形,1000以下有强峰。 910端基氢,再有一氢990。 顺式二氢690,反式移至970; 单氢出峰820,干扰顺式难确定。 炔氢伸展三千三,峰强很大峰形尖。 三键伸展二千二,炔氢摇摆六百八。 芳烃呼吸很特征,1600~1430。 1650~2000,取代方式区分明。 900~650,面外弯曲定芳氢。 五氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢。 醇酚羟基易缔合,三千三处有强峰。 C-O伸展吸收大,伯仲叔醇位不同。 1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。 1110醚链伸,注意排除酯酸醇。 若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。 苯环若有甲氧基,碳氢伸展2820。 次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。 缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。 羰基伸展一千七,2720定醛基。 吸电效应波数高,共轭则向低频移。 张力促使振动快,环外双键可类比。 二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。 羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。 1740酯羰基,何酸可看碳氧展。 1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。 1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。 羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。 伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。 胺尖常有干扰见,N-H伸展三千三, 叔胺无峰仲胺单,伯胺双峰小而尖。 1600碳氢弯,芳香仲胺千五偏。 八百左右面内摇,确定最好变成盐。 伸展弯曲互靠近,伯胺盐三千强峰宽, 仲胺盐、叔胺盐,2700上下可分辨, 亚胺盐,更可怜,2000左右才可见。 硝基伸缩吸收大,相连基团可弄清。 1350、1500,分为对称反对称。 氨基酸,成内盐,3100~2100峰形宽。 1600、1400酸根展,1630、1510碳氢弯。 盐酸盐,羧基显,钠盐蛋白三千三。 矿物组成杂而乱,振动光谱远红端。 钝盐类,较简单,吸收峰,少而宽。 注意羟基水和铵,先记几种普通盐。 1100是硫酸根,1380硝酸盐, 1450碳酸根,一千左右看磷酸。 硅酸盐,一峰宽,1000真壮观。 勤学苦练多实践,红外识谱不算难。
[font=黑体][size=4]红外识谱歌[/size][/font][font=楷体_GB2312]外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。[/font]
红外光谱议识谱歌外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
红外识谱歌外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。五氢吸收有两峰,700和750;四氢只有750,二氢相邻830;间二取代出三峰,700、780,880处孤立氢醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲,1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准,1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰,环氧乙烷有三峰,1260环振动,九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。酸酐也有C-O键,开链环酐有区别,开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽,920,钝峰显,羧基可定二聚酸、酸酐千八来偶合,双峰60严相隔,链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰,1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸,1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰;N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550;碳氮伸展酰胺III,1400强峰显。胺尖常有干扰见,N-H伸展三千三,叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽,仲胺盐、叔胺盐,2700上下可分辨,亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐,1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
红外可分远中近,中红特征指纹区, 1300来分界,注意横轴划分异。 看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。芳烃呼吸很特征,1600~1430。1650~2000,取代方式区分明。900~650,面外弯曲定芳氢。 氢吸收有两峰,700和750; 四氢只有750,二氢相邻830; 间二取代出三峰,700、780,880处孤立氢。 醇酚羟基易缔合,三千三处有强峰。C-O伸展吸收大,伯仲叔醇位不同。1050伯醇显,1100乃是仲, 1150叔醇在,1230才是酚。1110醚链伸,注意排除酯酸醇。若与π键紧相连,二个吸收要看准, 1050对称峰,1250反对称。苯环若有甲氧基,碳氢伸展2820。次甲基二氧连苯环,930处有强峰, 环氧乙烷有三峰,1260环振动, 九百上下反对称,八百左右最特征。缩醛酮,特殊醚,1110非缩酮。 酸酐也有C-O键,开链环酐有区别, 开链强宽一千一,环酐移至1250。羰基伸展一千七,2720定醛基。吸电效应波数高,共轭则向低频移。张力促使振动快,环外双键可类比。二千五到三千三,羧酸氢键峰形宽, 920,钝峰显,羧基可定二聚酸、 酸酐千八来偶合,双峰60严相隔, 链状酸酐高频强,环状酸酐高频弱。羧酸盐,偶合生,羰基伸缩出双峰, 1600反对称,1400对称峰。1740酯羰基,何酸可看碳氧展。1180甲酸酯,1190是丙酸, 1220乙酸酯,1250芳香酸。1600兔耳峰,常为邻苯二甲酸。 氮氢伸展三千四,每氢一峰很分明。羰基伸展酰胺I,1660有强峰; N-H变形酰胺II,1600分伯仲。伯胺频高易重叠,仲酰固态1550; 碳氮伸展酰胺III,1400强峰显。 胺尖常有干扰见,N-H伸展三千三, 叔胺无峰仲胺单,伯胺双峰小而尖。1600碳氢弯,芳香仲胺千五偏。八百左右面内摇,确定最好变成盐。伸展弯曲互靠近,伯胺盐三千强峰宽, 仲胺盐、叔胺盐,2700上下可分辨, 亚胺盐,更可怜,2000左右才可见。硝基伸缩吸收大,相连基团可弄清。 1350、1500,分为对称反对称。氨基酸,成内盐,3100~2100峰形宽。1600、1400酸根展,1630、1510碳氢弯。盐酸盐,羧基显,钠盐蛋白三千三。矿物组成杂而乱,振动光谱远红端。钝盐类,较简单,吸收峰,少而宽。注意羟基水和铵,先记几种普通盐。1100是硫酸根,1380硝酸盐, 1450碳酸根,一千左右看磷酸。硅酸盐,一峰宽,1000真壮观。勤学苦练多实践,红外识谱不算难。
范海福 中国科学院,物理研究所,北京,100080物质的各种宏观性质源出于本身的微观结构。探索物质结构与性质之间的关系,是凝聚态物理、结构化学、材料科学、分子生物等许多学科的一个重要研究内容。晶体结构分析,是在原子的层次上测定固态物质微观结构的主要手段,它与上述众多学科有着密切的联系。就其本身而言,晶体结构分析是物理学中的一个小分支。这主要研究如何利用晶态物质对X-射线、电子、以及中子的衍射效应来测定物质的微观结构。晶体结构分析服务于许多不同的学科,因而许多学科的发展都对晶体结构分析产生深刻的影响。另一方面,晶体结构分析有自己独立的体系,它本身的发展又对所服务的学科起着促进作用。 晶体结构分析是伦琴发现X-射线以后创站的最重要学科之一。它奠基于物理学的几项重要进展。其中包括1895年W. C. Roentgen发现X-射线,1912年M. von Laue发现晶体对X-射线的衍射,1927年C. J. Davisson和G. P. Thomson发现晶体对电子的衍射,以及1931年E. Ruska建造第一台电子显微镜。上述几项重大的物理学进展使人类掌握了在原子层次上研究物质内部结构的手段,它们分别获得1901、1914、1937和1986年的诺贝尔物理学奖。其中,1901年伦琴获得的诺贝尔奖还是历史上第一个诺贝尔物理奖。通过研究物质内部结构与性质的关系,晶体结构分析有力地促进了各相关学科的发展。晶体结构分析的发展,是一个不断完善自身和不断扩大应用的过程。诺贝尔将的年谱记录了晶体结构分析历史上的重大事件并展示了它与其他学科相互作用所产生的丰硕成果。 晶体结构分析的方法主要有两大类。这就是以X-射线衍射为代表的衍射分析方法和以电子显微术为代表的显微成像方法。电了显微成像也可以认为是两上相继的电子衍射过程。因此,可以说衍射分析是晶体结构分析的核心。用衍射分析方法测定晶体结构的理论依据,在于晶体结构同它的衍射效应之间存在着互为Fourier变换的关系。这里说的衍射效应,是指从晶体向各个方向发出的衍射的振幅和相位。从衍射实验可以记录下各个方向上衍射波的振幅。但是在目前以及可见的将来,还不容易找到有普遍意义的实用方法来记录由晶体发出的衍射波的相位。因此要想从衍射效应的Fourier变换解出晶体结构,必须先设法找回"丢失了的"相位。这就是晶体学中的"相位问题",它一直是研究晶体结构分析方法的关键问题。 紧接着Laue发现X-射线衍射,Bragg父子 (W. H. Bragg和W. L. Bragg) 就迅速建立了用X-射线衍射方法测定晶体结构的实验手段和理论基础。这使人类得以定量地观测原子在晶体中的位置。为此他们两人同获1915年的诺贝尔物理学奖。晶体结构分析最初用于一些简单的无机化合物。对碱金属卤化物结构的研究导至W. L. Bragg提出原子半径的概念。不久Bragg又将晶体结构分析应用于研究硅酸盐以及金属和合金。硅酸盐晶体结构分析的工作为硅酸盐结构化学提供了最早的实验基础,而有关金属和合金的工作则作物理冶金、金属物理、以及相平衡图的研究推上了一个新的台阶,使有关工作深入到原子的层次。 晶体结构分析在研究无机化合物上取得成功,引起人们对有机物尤其是生命物质内部结构的兴趣。英国从二十年代中期就开始研究有机物晶体结构。但是过了十年多仍未见有重大的突破。原因是当时的分析技术和方法还很原始。于是迎来了三、四十年代晶体结构分析方法和技术大发展的时期。如前所述,晶体结构分析中所谓"相位问题"。早期的晶体结构分析用以解决相位问题的方法是所谓尝试法。其要点是:先根据已尼掌握的线索猜想出一个结构模型,再从这个模型计算出相应的一组理论衍射强度,然后同实验所犁衍射强度作比较并据此对模型进行修改。。上述步骤须经多次反复,直至理论和实验的衍射强度得以吻合。用这样的"方法"来测定晶体结构,说明科学试验却更像艺术创作。它显然适应不了测定复杂的晶体结构的需要。早在二十年代后期,英国的W. L. Bragg和J. M. Cork为解决相位问题分别提出了所谓重原子法和同晶型置换法。重原子法的大意是:假定晶体中含有少数原子序较大的原子,即所谓重原子,而且它们的位置是已知的,这时就可以计算出重原子对相位的贡献并以此代替由全体原子贡献的相位。用这样的相位配以由实验测得的衍射振幅就可以近似地计算出一幅代表晶体结构的电子密度图。同晶型置找法的要点则是如果能够制备出待测晶体的重原子衍生物,而且衍生物的晶体与母体晶体是"同晶型"这时如果已知重原子的位置,就可以根据母体和衍生物两者在衍射强度上的差异来推算相应的衍射相位。这两种方法后来在一系列有机物以及蛋白质的晶体结构分析中作出了关键性的贡献。但是它们的诞生后相当长的一段时间里并未发挥很大的作用。原因是它们都依赖于已知的重原子位置而当时还没有便确定重原子位置的方法。1934年,美国的A. L. Patterson提出用衍射振幅的平方为系数以计算Fourier级数,从而绕开相位问题。Patterson指出,这样一个级数是晶体中电子密度分布函数的自卷积,在一定的条件下可以从中提取出有关晶体中原子位置,首先是重原子位置的信息。这个用衍射振幅平方计算的Fourier级数后来被称作Patterson函数,相应的分析方法称作Patterson法。经过几年发展之后,Patterson法和以它为基础的重原子法、同晶型置换法等就成了X-射线单晶体结构分析中用以处理相位问题最有效的手段。再加上实验技术和结构精修技术的改进,晶体结构分析达到了一个机关报的不平并终于打开了有机物和生命物质的大宝藏。 美国L. Pauling领导的小组花了十几年的时间,测定了一系列的氨基酸和肽的晶体结构,从中总结出形成多肽链构型的基本原则并在1951年推断多肽链将形成a-螺旋构型或折叠层构型。这是通过总结小分子结构规律预言生物大分子结构特征的非常成功的范例。为此Pauling获得1954年的诺贝尔化学奖。英国D. Hodgkin领导小组测定了一系列重要的生物化学物质的晶体结构,其中包括青酶素和维生素 。她因此获得1964年的诺贝尔化学奖。美国W. N. Lipscomb研究硼烷结构化学的工作获得1975年的诺贝尔化学奖。所有这些获奖工作都是以晶体结构分析为研究手段。可以说,没有晶体结构分析本身在理论和技术上的长期积累,就不会有上面几个诺贝尔奖。 英国的J. D. Bernal早在三十年代中期就开始用X-射线衍射研究蛋白质的结构。但是真正取得进展是在W. L. Bragg主持Cavendish实验室之后。这里还有一段插曲。原来在E. Rutherford主持下,英国剑桥大学的Cavendish实验室是国际上原子物理学的研究中心。随着学科的发展、国力的变化、加之剑桥大学本身的局限,及至1938年W. L. Bragg接任时Cavendish的地位已开始下降。Bragg上任后果断地顺应了形势,主动放弃了"原子物理国际中心"的地位,改而抓住当时物理学上的两项新应用:X-射线衍射分析用于生物以及雷达技术用于天文学。这一举措使英国得以在创建分子生物和射电天文学上"领导世界新潮流"。 分子生物学发展史上具有划时代意义的发现中,有两项出自Cavendish实验室。第一项是1953年J. D. Watson和F. H. C. Crick根据X-射线衍射实验建立了脱氧核糖核酸 (DNA) 的双螺旋结构。它把遗传学的研究推进到分子的水平。这项工作获得了1962年的诺贝尔生理学和医学奖。另一项是用X-射线衍射分析方法测定肌红蛋白和血红蛋白晶体结构的工作。它始于三十年代,前后延续了二十多年并牵涉到为数众多的科学家。这两个蛋白质的晶体结构终于在1960年被测定出来。这项工作不仅首次揭示了生物大分子内部的立体结构,还为测定生物大分子晶体结构提供了一种沿用至今的有效方法--多对同晶型置换法。它以原有的同晶型置换法为基础,但是在实验技术和分析理论上都加入了崭新的内容。作为这项工作的代表人物,J. C. Kendrew和M. F. Perutz获得1962年的诺贝尔化学奖。看到成就的辉煌,不由得也想起探索的艰辛:1947年,战后的英国,科研经费拮据。为了给正在从事蛋白质晶体结构分析的J. C. Kendrew和M. F. Perutz寻求资助,W. L. Bragg找到英国医学研究委员分 (MRC)。他告诉MRC的主管:"…如果能获得资助,我们的研究结果会有助于在分子层次上了解生命的运作。不过,即便如此,要想在医学上产生任何一点效益,大概还得有一段很长的时间"。MRC当时的主管承担了这一风险,建立了一个只包含Kendrew和Perutz两个人的MRC研究小组。这一慷慨的支持,过了十五年之后才开始得到回报。顺便说一句:那个MRC小组现在已经变成拥有上百名学者的、世界著名MRC分子生物学实验室。在Kendrew和Perutz两人之后由于测定蛋白质晶体结构而获诺贝尔奖的还有美国的J. Deisenhofer和德国的R. Huber和H. Michel。他们因测定了光合作用中心的三维结构而获得1988年诺贝尔化学奖。
物质的各种宏观性质源出于本身的微观结构。探索物质结构与性质之间的关系,是凝聚态物理、结构化学、材料科学、分子生物等许多学科的一个重要研究内容。晶体结构分析,是在原子的层次上测定固态物质微观结构的主要手段,它与上述众多学科有着密切的联系。就其本身而言,晶体结构分析是物理学中的一个小分支。这主要研究如何利用晶态物质对X-射线、电子、以及中子的衍射效应来测定物质的微观结构。晶体结构分析服务于许多不同的学科,因而许多学科的发展都对晶体结构分析产生深刻的影响。另一方面,晶体结构分析有自己独立的体系,它本身的发展又对所服务的学科起着促进作用。 晶体结构分析是伦琴发现X-射线以后创站的最重要学科之一。它奠基于物理学的几项重要进展。其中包括1895年W. C. Roentgen发现X-射线,1912年M. von Laue发现晶体对X-射线的衍射,1927年C. J. Davisson和G. P. Thomson发现晶体对电子的衍射,以及1931年E. Ruska建造第一台电子显微镜。上述几项重大的物理学进展使人类掌握了在原子层次上研究物质内部结构的手段,它们分别获得1901、1914、1937和1986年的诺贝尔物理学奖。其中,1901年伦琴获得的诺贝尔奖还是历史上第一个诺贝尔物理奖。通过研究物质内部结构与性质的关系,晶体结构分析有力地促进了各相关学科的发展。晶体结构分析的发展,是一个不断完善自身和不断扩大应用的过程。诺贝尔将的年谱记录了晶体结构分析历史上的重大事件并展示了它与其他学科相互作用所产生的丰硕成果。 晶体结构分析的方法主要有两大类。这就是以X-射线衍射为代表的衍射分析方法和以电子显微术为代表的显微成像方法。电了显微成像也可以认为是两上相继的电子衍射过程。因此,可以说衍射分析是晶体结构分析的核心。用衍射分析方法测定晶体结构的理论依据,在于晶体结构同它的衍射效应之间存在着互为Fourier变换的关系。这里说的衍射效应,是指从晶体向各个方向发出的衍射的振幅和相位。从衍射实验可以记录下各个方向上衍射波的振幅。但是在目前以及可见的将来,还不容易找到有普遍意义的实用方法来记录由晶体发出的衍射波的相位。因此要想从衍射效应的Fourier变换解出晶体结构,必须先设法找回"丢失了的"相位。这就是晶体学中的"相位问题",它一直是研究晶体结构分析方法的关键问题。 紧接着Laue发现X-射线衍射,Bragg父子 (W. H. Bragg和W. L. Bragg) 就迅速建立了用X-射线衍射方法测定晶体结构的实验手段和理论基础。这使人类得以定量地观测原子在晶体中的位置。为此他们两人同获1915年的诺贝尔物理学奖。晶体结构分析最初用于一些简单的无机化合物。对碱金属卤化物结构的研究导至W. L. Bragg提出原子半径的概念。不久Bragg又将晶体结构分析应用于研究硅酸盐以及金属和合金。硅酸盐晶体结构分析的工作为硅酸盐结构化学提供了最早的实验基础,而有关金属和合金的工作则作物理冶金、金属物理、以及相平衡图的研究推上了一个新的台阶,使有关工作深入到原子的层次。 晶体结构分析在研究无机化合物上取得成功,引起人们对有机物尤其是生命物质内部结构的兴趣。英国从二十年代中期就开始研究有机物晶体结构。但是过了十年多仍未见有重大的突破。原因是当时的分析技术和方法还很原始。于是迎来了三、四十年代晶体结构分析方法和技术大发展的时期。如前所述,晶体结构分析中所谓"相位问题"。早期的晶体结构分析用以解决相位问题的方法是所谓尝试法。其要点是:先根据已尼掌握的线索猜想出一个结构模型,再从这个模型计算出相应的一组理论衍射强度,然后同实验所犁衍射强度作比较并据此对模型进行修改。。上述步骤须经多次反复,直至理论和实验的衍射强度得以吻合。用这样的"方法"来测定晶体结构,说明科学试验却更像艺术创作。它显然适应不了测定复杂的晶体结构的需要。早在二十年代后期,英国的W. L. Bragg和J. M. Cork为解决相位问题分别提出了所谓重原子法和同晶型置换法。重原子法的大意是:假定晶体中含有少数原子序较大的原子,即所谓重原子,而且它们的位置是已知的,这时就可以计算出重原子对相位的贡献并以此代替由全体原子贡献的相位。用这样的相位配以由实验测得的衍射振幅就可以近似地计算出一幅代表晶体结构的电子密度图。同晶型置找法的要点则是如果能够制备出待测晶体的重原子衍生物,而且衍生物的晶体与母体晶体是"同晶型"这时如果已知重原子的位置,就可以根据母体和衍生物两者在衍射强度上的差异来推算相应的衍射相位。这两种方法后来在一系列有机物以及蛋白质的晶体结构分析中作出了关键性的贡献。但是它们的诞生后相当长的一段时间里并未发挥很大的作用。原因是它们都依赖于已知的重原子位置而当时还没有便确定重原子位置的方法。1934年,美国的A. L. Patterson提出用衍射振幅的平方为系数以计算Fourier级数,从而绕开相位问题。Patterson指出,这样一个级数是晶体中电子密度分布函数的自卷积,在一定的条件下可以从中提取出有关晶体中原子位置,首先是重原子位置的信息。这个用衍射振幅平方计算的Fourier级数后来被称作Patterson函数,相应的分析方法称作Patterson法。经过几年发展之后,Patterson法和以它为基础的重原子法、同晶型置换法等就成了X-射线单晶体结构分析中用以处理相位问题最有效的手段。再加上实验技术和结构精修技术的改进,晶体结构分析达到了一个机关报的不平并终于打开了有机物和生命物质的大宝藏。 美国L. Pauling领导的小组花了十几年的时间,测定了一系列的氨基酸和肽的晶体结构,从中总结出形成多肽链构型的基本原则并在1951年推断多肽链将形成a-螺旋构型或折叠层构型。这是通过总结小分子结构规律预言生物大分子结构特征的非常成功的范例。为此Pauling获得1954年的诺贝尔化学奖。英国D. Hodgkin领导小组测定了一系列重要的生物化学物质的晶体结构,其中包括青酶素和维生素 。她因此获得1964年的诺贝尔化学奖。美国W. N. Lipscomb研究硼烷结构化学的工作获得1975年的诺贝尔化学奖。所有这些获奖工作都是以晶体结构分析为研究手段。可以说,没有晶体结构分析本身在理论和技术上的长期积累,就不会有上面几个诺贝尔奖。英国的J. D. Bernal早在三十年代中期就开始用X-射线衍射研究蛋白质的结构。但是真正取得进展是在W. L. Bragg主持Cavendish实验室之后。这里还有一段插曲。原来在E. Rutherford主持下,英国剑桥大学的Cavendish实验室是国际上原子物理学的研究中心。随着学科的发展、国力的变化、加之剑桥大学本身的局限,及至1938年W. L. Bragg接任时Cavendish的地位已开始下降。Bragg上任后果断地顺应了形势,主动放弃了"原子物理国际中心"的地位,改而抓住当时物理学上的两项新应用:X-射线衍射分析用于生物以及雷达技术用于天文学。这一举措使英国得以在创建分子生物和射电天文学上"领导世界新潮流"。 分子生物学发展史上具有划时代意义的发现中,有两项出自Cavendish实验室。第一项是1953年J. D. Watson和F. H. C. Crick根据X-射线衍射实验建立了脱氧核糖核酸 (DNA) 的双螺旋结构。它把遗传学的研究推进到分子的水平。这项工作获得了1962年的诺贝尔生理学和医学奖。另一项是用X-射线衍射分析方法测定肌红蛋白和血红蛋白晶体结构的工作。它始于三十年代,前后延续了二十多年并牵涉到为数众多的科学家。这两个蛋白质的晶体结构终于在1960年被测定出来。这项工作不仅首次揭示了生物大分子内部的立体结构,还为测定生物大分子晶体结构提供了一种沿用至今的有效方法--多对同晶型置换法。它以原有的同晶型置换法为基础,但是在实验技术和分析理论上都加入了崭新的内容。作为这项工作的代表人物,J. C. Kendrew和M. F. Perutz获得1962年的诺贝尔化学奖。看到成就的辉煌,不由得也想起探索的艰辛:1947年,战后的英国,科研经费拮据。为了给正在从事蛋白质晶体结构分析的J. C. Kendrew和M. F. Perutz寻求资助,W. L. Bragg找到英国医学研究委员分 (MRC)。他告诉MRC的主管:"…如果能获得资助,我们的研究结果会有助于在分子层次上了解生命的运作。不过,即便如此,要想在医学上产生任何一点效益,大概还得有一段很长的时间"。MRC当时的主管承担了这一风险,建立了一个只包含Kendrew和Perutz两个人的MRC研究小组。这一慷慨的支持,过了十五年之后才开始得到回报。顺便说一句:那个MRC小组现在已经变成拥有上百名学者的、世界著名MRC分子生物学实验室。在Kendrew和Perutz两人之后由于测定蛋白质晶体结构而获诺贝尔奖的还有美国的J. Deisenhofer和德国的R. Huber和H. Michel。他们因测定了光合作用中心的三维结构而获得1988年诺贝尔化学奖。
摘要:通过试验和观察,研究了活性污泥中丝状菌与絮体结构的关系。常见的活性污泥絮体可分为六大类型,在不同的处理工艺和运行条件下,各类型污泥比例不同,丝状菌在污泥絮体的形成过程中所起的作用也不相同。而在活性污泥膨胀时,生物相结构中的丝状菌可分为结构性的和非结构性的两大类,它们起着不同的作用,运行中必须通过不同的方法和措施加以防治。丝状微生物是一大类菌体相连而形成丝状的微生物的统称,其中包括丝状细菌、丝状真菌、丝状藻类等[1]。荷兰学者Eikelboom将丝状微生物分为29个类型、7个群,并制成了活性污泥丝状微生物检索表。 丝状微生物的功能与结构形态密切相关,长丝状形态有利于其在固相上附着生长,保持一定的细胞密度,防止单个细胞状态时被微型动物吞食;细丝状形态的比表面积大,有利于摄取低浓度底物,在底物浓度相对较低的条件下比胶团菌增殖速度快,在底物浓度较高时则比胶团菌增殖速度慢。许多丝状微生物表面具有胶质的鞘,能分泌粘液,粘液层能够保证一定的胞外酶浓度,并减少水流对细胞的冲刷,其中还含有特定的抗体,以防止其他生物附着。 丝状微生物种类繁多,对生长环境要求低。其本身生理生长特性很特别:增殖速率快、吸附能力强、耐供氧不足能力以及在低基质浓度条件下的生活能力都很强,因此在废水生物处理生态系统中存活的种类多,数量大。如何使丝状微生物相互聚集,使之在废水处理中达到较好的泥水分离效果,如何确定丝状微生物同其他微生物的相互作用,以及不同丝状微生物的最适需氧量等,都是需要进一步研究的问题。1 试验设计及过程试验分别在本院给水排水实验室、重庆市唐家桥污水处理厂、重庆市渝北区城南污水处理厂进行。活性污泥采样自本实验室活性污泥法小试反应器、唐家桥污水处理厂和城南污水处理厂的曝气池、初沉池和二沉池。通过镜检观察记录活性污泥絮体大小、形态和结构,对不同反应器的丝状微生物进行鉴定,从而寻找丝状微生物与絮体形态结构之间的关系。试验历时5个月。 丝状微生物鉴定采用Eikelboom法,镜检观察以下八项特征:①是否存在衣鞘;②滑行运动;③真、假分枝;④丝状体长度、形状、性质;⑤细胞直径、长度、性质;⑥革兰氏染色反应;⑦纳氏染色反应;⑧有无胞含体(聚-β-羟基丁酸PHB、硫粒、多聚磷酸盐等)。染色采用石炭酸复红染色法、革兰氏染色法、纳氏染色法和积硫试验法。通过目微尺测定污泥絮体直径,记录各种大小、形状和结构的絮体数量,归纳污泥絮体的主要类型及特征。通过大量观察,寻找丝状微生物种类、浓度与污泥絮体大小、形状、结构的关系。2 试验结果2.1 絮体结构形态类型 通过大量的观察发现,活性污泥在正常运行和膨胀时呈现不同的结构形态和种类。正常运行时活性污泥结构形态可分为四类,Ⅰ型:致密、细小,看不到丝状菌为骨架的污泥;Ⅱ型:有明显丝状骨架、呈长条形的污泥;Ⅲ型:厚实、具有网状结构的巨型污泥;Ⅳ型:有孔洞结构的巨型污泥。污泥膨胀时其结构形态可分为两类,Ⅴ型:结构丝状菌大量生长、伸长,絮体结构松散;Ⅵ型:非结构丝状菌大量生长,不形成絮体。 试验过程中发现,Ⅰ型污泥在两污水厂正常运行的曝气池中所占比例较低,城南污水厂为10%左右,唐家桥污水厂更低,而在二沉池上清液中比例较高,因此它是从良好结构的污泥上脱落下来的,在二沉池随出水流失。正常运行时长条形污泥、网状污泥和孔洞污泥(Ⅱ、Ⅲ、Ⅳ型)占很高比例,两污水厂中均占90%以上。根据絮体伸出的部分丝状菌,可以判断这些具有良好结构的污泥是以丝状菌为骨架,胶团菌附着于其上而形成的。它们是去除有机物的主要部分。 在混合液中可见到其他丝状微生物游离于菌胶团之外,见不到附着生长物,三种样本见到的菌种有:球衣菌、发硫菌、0803型、0581型、硬发菌、链球菌等,但数量都十分少。 试验过程中,城南污水厂由于发生停电事故时仍保持进水流量,发生了结构丝状菌大量增殖的现象,污泥结构呈松散状(Ⅴ型),SVI达到142mL/g干污泥;待供电正常,按正常方式运行一段时间后,污泥结构恢复正常,SVI回落至90mL/g 干污泥。而活性污泥小试过程中多次出现污泥膨胀,泥水分离困难(Ⅵ型),SVI高达500mL/g 干污泥以上,调节运行方式仍不能控制,镜检发现球衣菌、发硫菌大量增殖,最终通过投加漂白粉杀生剂再经逐步培养才恢复正常。2.2 微生物鉴定结果 根据Eikelboom法对作为污泥良好结构骨架的丝状菌进行鉴定,发现各处取样污泥的结构丝状菌特征一致:丝状体直径1.5~2μm,丝体长200μm左右,不运动,略弯,在絮体内扭曲,细胞呈柱状,长0.5~4μm,直径0.7~1.0μm,有鞘,横隔明显,常见分枝,有大量附着生长物,无硫粒,革兰氏染色阴性,纳氏染色可见兰灰色颗粒,呈阳性。 查丝状微生物鉴定表,找不到特征完全相符的种,比较接近的是Eikelboom1701型。Eikelboom1701的特征是:链状圆柱形细胞,被鞘紧裹,丝体长100~200μm,偶尔超过200μm,虽然丝体正常时稍弯,但可有很强的盘绕性,细胞长2.5~3.5μm,直径0.5~0.9μm,有鞘,有时可见PHB黑色小颗粒,横隔和缩缢明显,偶有假分枝,常有大量附着生长物,无硫粒,革兰氏染色阴性,纳氏染色阳性。3 分析与讨论3.1 絮体形成过程 许多絮体可以同时具有Ⅱ型、Ⅲ型、Ⅳ型污泥的多种特征,在絮体中心部分为孔洞结构,向四周伸展的长条形污泥相互搭接形成网状结构,最外侧则可见新伸出的骨架丝状菌。从这种污泥的形态可以推断其形成过程为:结构丝状菌交织生长,胶团菌附着其上形成新生污泥,新生污泥逐渐成熟形成条状、网状污泥,在氧和营养物充足等条件下,网状污泥的胶团菌增粗,网孔逐渐变小形成孔洞状,最后孔洞被填实,而结构丝状菌的伸出为胶团菌提供了新的附着面,包裹形成新的条状污泥,条状污泥相互交织又形成新的网状污泥,重复上述过程,形成更大的污泥絮体。 一些污泥能见到成节的形态,大的孔洞结构污泥之间由细的条状污泥连接,有的由丝状微生物连接,这种污泥的形成可能是絮体成长到一定成熟度后,由于内部供氧不足,促进了包埋于其中的结构丝状菌的生长,将絮体撑开导致结构松散形成节状。 还有极少量的污泥,可以见到极粗大的丝状骨架,上面附着胶团菌,经多次对比鉴定,这些丝状骨架为死亡累枝虫的杆,由于结构松散,这类污泥易于在二沉池发生漂浮,因此保持原生动物稳定的生长条件可以有效地减少二沉池的污泥上浮。3.2 丝状微生物与微生态群落的关系 试验表明,胶团菌与结构丝状菌之间相互依存,丝状微生物形成了絮体骨架,为絮体形成较大颗粒同时保持一定的松散度提供了必要条件。而胶团菌的附着使絮体具有一定的沉降性而不易被出水带走,并且由于胶团菌的包附使得结构丝状菌获得更加稳定、良好的生态条件,所以这两大类微生物在活性污泥中形成了特殊的共生体。 根据生态学的观点,环境因子对微生物个体的影响首先是影响某些敏感生物,然后通过微生物之间的相互作用逐步传递,最终当影响超过一定限度时引起结构上的波动。正是因为生态系统中生物种类多,并按一定结构组成了微生态群落,环境压力在逐级传递过程中受到消减,所以生态系统具备了一定抗冲击负荷的能力。与纯培养相比,生态系统能通过优势种群的变化维持良好的结构,而纯培养只需轻微刺激就会引起强烈反应,直接破坏其脆弱的结构。这也是保证活性污泥微生态群落稳定性的根本原因。 根据本试验结果,可以将活性污泥微生态群落描述如下:活性污泥微生态群
请问大师们研究拉伸对高分子、聚合物和纤维的结构性能(包括结晶)的影响我想找一下外文和中文的文献看看请问一下各位(1)国内的哪些专家在做这个方面或者相关的研究(2)国外哪些专家在做这个方面的研究(3)哪个国外期刊对这个方面的研究的报道比较牛一点因为我知道了专家的名字,找相关的文献要方便的多有知道的哥们和大师们帮个忙,告诉我一声。 谢谢!
请问各位:我打算开一个公司做钢结构无损检测(第三方),想申请CMA,CMA 的申请条件大概弄清楚了,但是今天刚刚发现,申请钢结构无损检测这个经营范围似乎还和省住房和城乡建设厅有瓜葛(附件是朋友给我的材料,这个检测资质貌似还要受省住房和城乡建设厅审核制约),这个省住房和城乡建设厅的要求(注册资本,固定资产,持证人员数量和证书等级等等都比申请CMA的入门要求提高很多)。那么问题来了:申请钢结构第三方无损检测作为公司主营业务,是不是一定要取得省住房和城乡建设厅的批准后才能开始CMA的申请认证? 没有省住房和城乡建设厅的批准可以直接申请CMA吗? 省住房和城乡建设厅绕不过去吗?
外可分远中近,中红特征指纹区,1300来分界,注意横轴划分异。看图要知红外仪,弄清物态液固气。样品来源制样法,物化性能多联系。 识图先学饱和烃,三千以下看峰形。2960、2870是甲基,2930、2850亚甲峰。1470碳氢弯,1380甲基显。二个甲基同一碳,1380分二半。面内摇摆720,长链亚甲亦可辨。烯氢伸展过三千,排除倍频和卤烷。末端烯烃此峰强,只有一氢不明显。化合物,又键偏,~1650会出现。烯氢面外易变形,1000以下有强峰。910端基氢,再有一氢990。顺式二氢690,反式移至970;单氢出峰820,干扰顺式难确定。炔氢伸展三千三,峰强很大峰形尖。三键伸展二千二,炔氢摇摆六百八。
绪 论第一节 土壤在人类农业和自然环境中的作用一、土壤是人类农业生产的基地“民以食为天,食以土为本”概括了人类—农业—土壤之间的关系。农业是人类生存的基础,而土壤是农业的基础。一、 (一)土壤是植物生长繁育和生物生产的基地农业生产的特点是生产出有生命的生物有机体,其中最基本的任务是发展植物赖以生存的绿色植物的生产。绿色植物的生长发育需要五个要素:光、热、空气(氧气和二氧化碳)、水分和养分。其中养分和水分通过根系从土壤中吸取。植物能立足于自然界,依靠根系伸展在土壤中,获得土壤的机械支撑,才不至于倒下。1、营养库的作用 植物需要的营养元素除主要来自空气外,及中量元素、微量元素和水分则主要来自土壤。2、养分转化和循环作用 无机物通过生物作用转为有机物(光合作用),有机物质经过矿质化作用变无机物,此过程在土壤中完成,3、雨水涵养作用 土壤是地球陆地表面具有生物活性和多孔结构的物质,具有很强的吸水和持水能力。据统计,地球上的淡水总贮(zhu )水量为0.39亿㎞3,其中被冰雪封存和埋藏在地壳深层的水有0.349亿㎞3。可供人类生产和生活的循环淡水总贮量只有0.041亿㎞3,仅占总淡水量的10.5﹪。在0.041亿㎞3的循环淡水中,循环地下水占95.12﹪湖泊水占2.95﹪,土壤水占1.59﹪,江河水占0.03﹪,大气水0.34﹪,土壤贮水量明显大于江河和大气的贮水量。土壤的雨水涵养功能与土壤的总孔度、有机质含量等土壤的理化性质和植被覆盖度有密切关系。4、生物的支撑作用 土壤不仅提供给植物的营养物质,还是植物根系的伸展和穿插,获得机械支撑的物质环境。5、稳定和缓冲环境变化的作用土壤处于大气圈、水圈、岩圈及生物圈的交界面,是地球表面各种物理、化学和生物化学过程的反应界面,是物质和能量交换、迁移等过程最复杂,最频繁的地带。
目前,单晶硅太阳电池的输出电压约为0.6V左右,其最大输出的功率和 太阳电池本身的效率与表面积有关。譬如说一个效率16%、6时的太阳电 池,最大输出功率约为2.5W。一个太阳电池输出电压和输出功率对大部分的 电器产品相对偏低,要和一般用电兼容或配合应用,就将多个太阳电池并联和串联起来形成模块(module),其中串联的功用,是为了提高输出电压,而并联的功用,是为了增加输出功率。同样的道理,若需要再提高模块的输出电压或 输出功率,多个模块并联或串联起来就形成数组(array)系统。而一般太阳能应 用系统(system),不仅只有电池、模块、或数组,还可能包括储电装置(storage devices)、功率调 器(power conditioner)、和安装固定结构(mounting structures),这些接口设备,统称为平衡系统(balance of systems)。下面,我们 就简单地介绍太阳电池的基本结构,了解太阳电池工作原理、制造程序,包括半导体材料。 在不同的材料和制造工艺程序下,会产生不同结构的太阳电池。但归纳而言,太阳电池最基本的结构可分为基板、p-n二极管、抗反射层、和金属电极 四个主要部分。基板(substrate)是太阳电池的主体,p-n二极管是光生伏特效应 的来源,抗反射层乃在减少入射光的反射来增强光电流,金属电极则是连接器件和外部负载。 所谓ingot-based的太阳电池是使用芯片(wafer) 当基板,芯片本身就是光生伏特的作用区。因为是用芯片作基板,一般就使用扩散(diffusion)工艺技 术,在p-型芯片上进行n-型扩散,或在n-型芯片上进行p-型扩散,形成p-n二极管。单晶娃和多晶硅太阳电池都是ingot-based,其芯片是由硅ingot切割而 得。工业界使用的太阳电池硅芯片,大都是p型。当然硅芯片的制造,不一定 非由ingot切割不可,也有其它特殊的方式,如ribbon或sheet制造方式。 薄膜太阳电池则可以使用玻璃、塑料、陶瓷、石墨,金属片等不同材料当基板,非晶或多晶薄膜光生伏特器件则沉积在基板上,基板本身并不参与光生 伏特作用。在薄膜太阳电池制造上,可使用各式各样的沉积技术,一层又一层 地把P-型或n-型材料沉积上去。常见的薄膜太阳电池有非晶硅、CUInSe2 (CIS)、CuInGaSe2 (CIGS)、和CdTe薄膜。随着薄膜技术的发展, microcrystalline,甚至nanocrytalline硅薄膜也被研究开发。薄膜太阳电池大优点就是生产成本较低,但其效率和稳定性较差。 III-V族(如GaAs、InP、GaN)太阳电池,则是使用不同的外延(epitaxy) 技术,如 metal-organic chemical vapor deposition (MOCVD),或 molecular beam epitaxy (MBE)方法,将p-型和n-型晶体直接长在芯片基板上,而基板本身通 常也不参与光生伏特作用。这样的epitaxy方式生长晶体的优点,使得电池结构 多样化,例如:异质结、多结、量子井、量子点、和超晶格等结构。正因如此,III-V族太阳电池通常具有较高的效率,但其生产成本也相对的偏高。 太阳电池的光照面一般都会有抗反射层或texture结构,来减少入射阳光的反射。如果没有的话,入射阳光会有约30%的反射损失,这对太阳电池而言足是相当严重的。晶硅太阳电池一般是使用氮化硅(SiN)来形成抗反射层,它不仅 能有效地减少入射光的反射,而且还有钝化(passivation)的作用,甚至能保 护太阳电池,有防刮伤、防湿气等功能。除了使用抗反射层外,一般单晶硅太 阳能电池,期光照的表面都会先经过texture处理,来更进一步地减少入射阳光 的反射。这个texture处理,会在表面形成大小不等的金字塔(pyramid)结构, 让入射光至少要经过芯片表面的二次反射,因此就大大地降低了入射光经过第 一次反射就折冋的几率。需要注意的是,因texture金字塔的大小约儿个um, 而一般n-型扩散的深度只有0.5um作用,所以二极管实际上是形成于textur金字塔的表面。[url=http://www.hyxyyq.com][color=#ffffff]手持万用表[/color][/url][url=http://www.hyxyyq.com][color=#ffffff]http://www.hyxyyq.com[/color][/url] 太阳电池需要金属电极一层来连接外部的电路。通常,光入射的表面有二条平行条状金属电极来提供外界连接的焊接处。背表面通常会全部涂上一层所谓的back surface field (BSF)金属层,在光入射的表面,会从条状金属电极,伸展出 一列很细的金属手指(finger)。BSF金属层可以增加载流子的收集,还可回收没 有被吸收的光子。金属finger的设计,除了要能有效地收集载流子,而且要尽 量减少金属线遮蔽入射光的比例,因光照面的金属线通常会遮蔽3〜 5%的入射 光。太阳电池金属电极用的材料通常是铝和其它金属的合金,但在薄膜太阳电池中,为了实现一体成型(monolithically)的要求,上层金属电极则会使用透明导电的氧化物 transparent conducting oxide (TCO)。 必须注意的是,有别于一般平板(flat plate)模块的结构,太阳电池还可以 使用额外的聚光器(concentrator)来增加入射光的强度。聚光器可以是一般透 镜,或是特殊结构的Fresne透镜,或者甚至是Fresnel zone plate。聚光器的使 用,可以大幅度地提高系统光照的有效面积。但是,聚光器要求太阳电池的正 射,因此应用上必须配合tracking系统。
有奖问答:织物组织结构与拉伸断裂强力有关,当其它条件相同时,就三原组织来说,强力最高的是 ?
急弹性,缓弹性和塑性三种变形的特点是怎样的? 急弹性变形的特征为外力作用时立即变形,外力去除则立即恢复。 缓弹性变形的特征为外力作用时变形逐渐增加,外力去除后逐渐恢复。 塑性变形的特征为外力作用时产生变形,外力去除后变形不恢复。 急弹性变形的实质为大分子链键长和链角的开合,皱曲大分子的部分伸展。 缓弹性变形的实质为大分子链屈曲伸展,滑移错位。 塑性变形的实质为大分子键质心不可恢复的粘性流动。







 我要推广仪器
我要推广仪器
 下载APP
下载APP