
定量蛋白质组学揭示内质网应激作用下蛋白质的构象变化


导读:本文通过设计一种半胱氨酸靶向探针,定量半胱氨酸残基的暴露率,系统地研究了蛋白质的结构以及结构的变化。
大家好,本周为大家分享一篇发表在Analytical Chemistry上的文章Quantitative Structural Proteomics Unveils the Conformational Changes of Proteins under the Endoplasmic Reticulum Stress1,文章的通讯作者是来自美国佐治亚理工学院的Ronghu Wu助理教授。在真核细胞中,内质网(endoplasmic reticulum,ER)负责蛋白质组中40%蛋白质的合成和成熟。蛋白质合成或折叠过程中的变化都将影响内质网的稳态,进而导致未折叠蛋白的积累和蛋白分泌效率的降低。在过去几十年的研究中,内质网应激反应被广泛研究,但是内质网应激反应后蛋白质折叠状态的变化却没有被深入研究。基于丰度的蛋白质组学方法不能直接用于分析蛋白质状态的变化,在这篇文章中,作者整合了半胱氨酸(cysteine,Cys)共价标记、选择性富集和定量蛋白质组学,称为半胱氨酸靶向共价蛋白绘制(cysteine targeted covalent protein painting,Cys-CPP),用于研究蛋白质组范围内的蛋白质结构和变化(图1A)。
使用CPP分析蛋白质结构,需要一种具有高反应活性的探针。作者设计了一种针对半胱氨酸的探针,其中包含半胱氨酸反应基团、用于富集的生物素部分和用于生成半胱氨酸特异性识别位点标签的可裂解连接部分(图1B)。以变性处理后的蛋白样品作为蛋白质展开形式的参考,计算肽段在原始样本和变性样本中的比例从而获得宝贵的蛋白质结构信息。
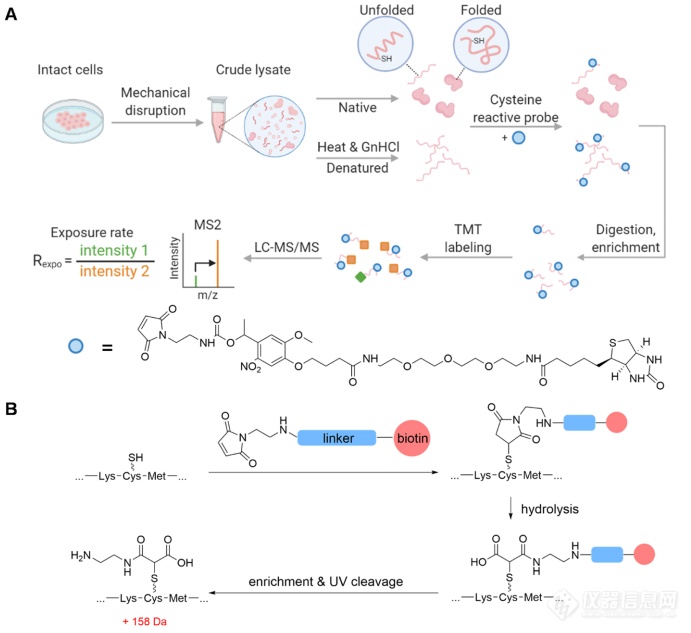
图1.利用半胱氨酸反应探针定量分析人细胞蛋白质组中半胱氨酸暴露率的原理。(A)Cys-CPP的一般工作流程。(B)半胱氨酸残基与探针之间的反应。富集后,进行紫外裂解,在修饰的半胱氨酸上留下一个小标记,用质谱进行位点特异性分析。
半胱氨酸暴露率Rexpo通过每条肽段在原始样本和变性样本中的比值进行计算。结果显示:(1)半胱氨酸的暴露率和溶剂可及性呈现正相关(图2C);(2)在丝氨酸和苏氨酸等极性氨基酸残基旁边的半胱氨酸具有相对较高的暴露率,这与人们普遍认为亲水残基更有可能暴露在蛋白质表面的观点一致;(3)甘氨酸和脯氨酸附近的半胱氨酸具有更高的暴露率,这是因为这两种氨基酸通常出现在蛋白质的转角和环结构中,对半胱氨酸的空间位阻较小;(4)半胱氨酸暴露率与其有/无序区(图2D)或所处二级结构(图2E)的相关性分析均表明,较低的暴露率与更稳定和结构化的局部环境有很好的相关性。这些数据结果共同证明目前的方法可以准确地测得半胱氨酸暴露率,并为蛋白质结构提供有价值的信息。
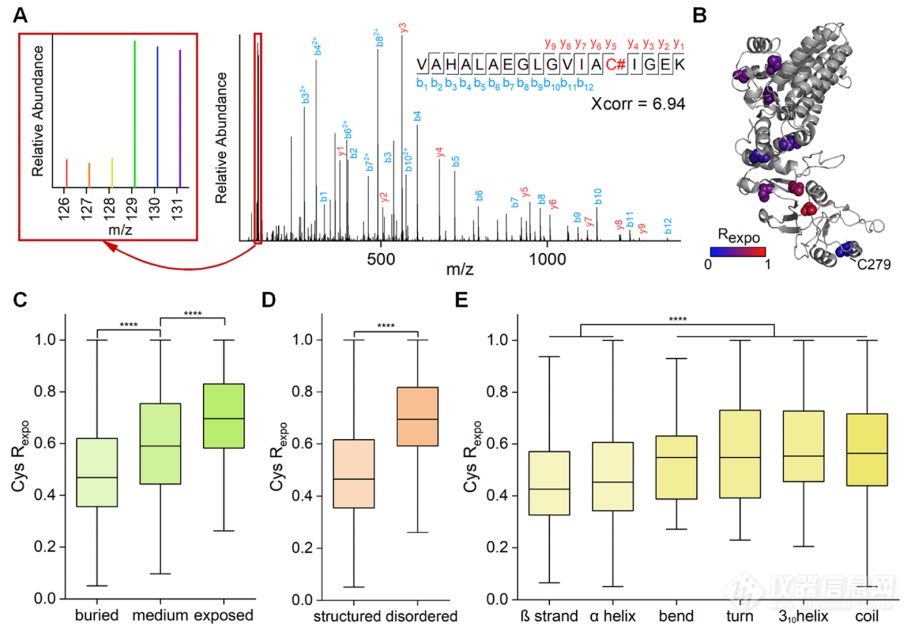
图2.HEK293T细胞中半胱氨酸暴露率的分析。(A) VAHALAEGLGVIAC#IGEK(#代表标记位点)的串联质谱样本。报告离子的强度使我们可以准确定量一个半胱氨酸的暴露率(左框为报告离子强度的放大视图)。(B)蛋白CCT3中被定量半胱氨酸的定位和暴露率演示(PDB代码:6qb8)。(C−E)比较不同的溶剂可及性(C)、预测无序区(D)和二级结构(E)的半胱氨酸暴露率。
衣霉素(Tunicamycin,Tm)可抑制 N-糖基化并阻断 GlcNAc 磷酸转移酶 (GPT)。由于蛋白质的N-糖基化经常发生在共翻译过程中,在蛋白质折叠的调节中起着至关重要的作用,所以衣霉素会引起细胞内质网中未折叠蛋白的积累并诱导内质网应激。基于此,作者用衣霉素对细胞进行处理,计算并对比了衣霉素处理样本和正常样本中的半胱氨酸暴露率。正如预期的那样,Tm处理样本中许多半胱氨酸的暴露率升高,且Tm对于蛋白质不稳定区域的作用尤为显著。根据Tm处理样本和正常样本之间半胱氨酸暴露率的差值,作者将所有位点划分为5个部分,在Tm处理下,近三分之一的半胱氨酸定位区域没有明显的结构变化(差值在-0.05~0.05之间),而28%的位点则高度暴露(差值>0.15)(图3B)。对这两种蛋白质进行基因本体(GeneOntology,GO)功能富集分析(图3C),结果显示:差值在-0.05~0.05之间的蛋白通常是糖异生或折叠过后具有良好结构区域的蛋白,而差值>0.15的蛋白则是与囊泡转运相关的蛋白。这表明抑制N-糖基化主要影响经典分泌途径中的蛋白质,与预期相符。
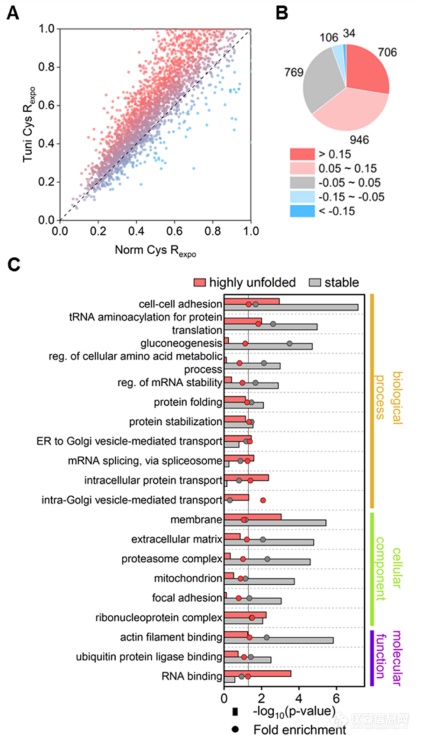
图3.利用Tm抑制蛋白质N-糖基化对蛋白质折叠影响的系统研究。(A)Tm处理和对照样品之间半胱氨酸暴露率的比较。(B) 不同暴露率变化范围内的蛋白质数量。(C)在具有高度展开或稳定区域半胱氨酸的蛋白之间进行GO功能富集分析。
由于Tm对于预先存在的、折叠良好的蛋白质所产生的影响可能远小于对新合成蛋白的影响,分别研究Tm对这两种蛋白的影响是必要的。作者通过将目前的方法Cys-CPP与细胞培养中氨基酸的稳定同位素标记(pSILAC)结合(图4A),探究了细胞中已存在蛋白和新合成蛋白在内质网应激作用下的不同变化。结果显示:(1)抑制N-糖基化对新合成蛋白的去折叠影响比对已存在蛋白的影响更显著(图4C);(2)N-糖基化除了调节蛋白质的二级结构外,在蛋白质三级或四级结构的形成中起着更重要的作用(图4D)。
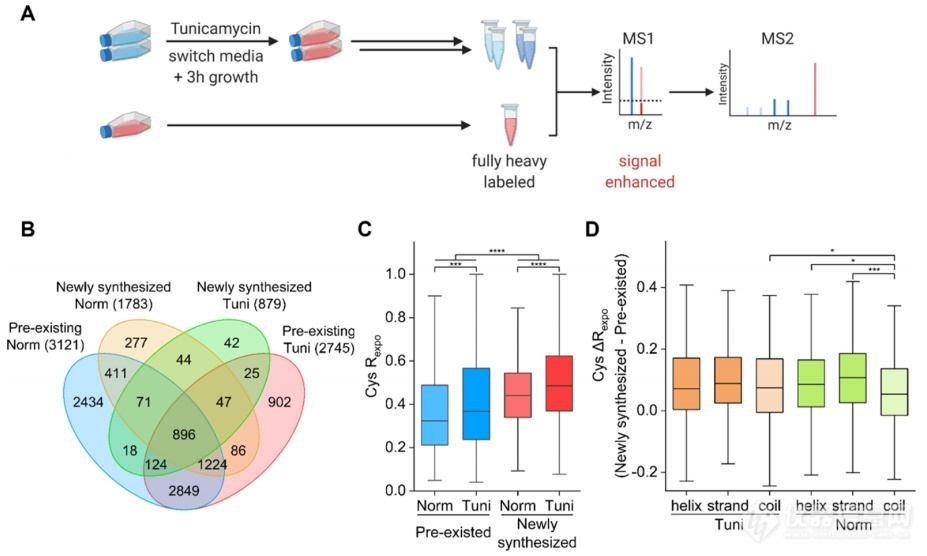
图4. 抑制N-糖基化对新合成蛋白和已存在蛋白折叠状态影响的研究。(A)量化新合成蛋白和已存在蛋白折叠状态变化的实验设置。(B) 经Tm处理和未经处理的细胞中新合成和已存在蛋白质的重叠。括号内为每组蛋白质数。(C)不同蛋白质组中暴露率的分布。(D) 在有或没有Tm处理的细胞中、在不同的二级结构下,新合成和已存在蛋白之间半胱氨酸暴露率的差值分布。
本文通过设计一种半胱氨酸靶向探针,定量半胱氨酸残基的暴露率,系统地研究了蛋白质的结构以及结构的变化。结果表明,半胱氨酸暴露率与蛋白质局部结构的相关性非常好。利用该方法,作者研究了Tm引起的内质网应激反应下细胞中蛋白质的结构变化。此外,通过将Cys-CPP与pSILAC结合,研究了在内质网应激反应下原有蛋白和新合成蛋白的结构变化差异,并详细分析了内质网应激对蛋白质去折叠的影响,深入和准确地了解内质网应激下的蛋白质结构变化,有助于深入了解蛋白质的功能和细胞活性。
参考文献:[1] Yin K, Tong M, Sun F, et al. Quantitative Structural Proteomics Unveil the Conformational Changes of Proteins under the Endoplasmic Reticulum Stress[J]. Analytical Chemistry, 2022,
来源于:仪器信息网













热门评论
最新资讯
新闻专题
更多推荐
