
红外多光子解离用于Top-Down表征膜蛋白复合物和G蛋白偶联受体


导读:作者证明了可以在改进的Orbitrap Eclipse质谱仪的高压QLIT下,通过红外照射可以完全释放一系列去垢剂胶束中的膜蛋白。
大家好,本周为大家分享一篇来自Angewandte Chemie - International Edition的文章:Infrared Multiphoton Dissociation Enables Top-Down Characterization of Membrane Protein Complexes and G ProteinCoupled Receptors[1],文章的通讯作者是牛津大学化学系的Carol V. Robinson教授。
非变性质谱(Native MS)是结构生物学中一个成熟的工具。在电喷雾电离过程中使用非变性缓冲液可以保存多组分蛋白质复合物之间的非共价相互作用,以及它们的配体、辅因子或其他结合蛋白。它可以用于探究蛋白质复合物的相互作用和功能,因为结合事件导致质量变化,可以在质谱仪中跟踪和剖析。然而,由于膜蛋白的疏水性,使得它们在传统的非变性质谱缓冲液中不溶且容易聚集,因此在非变性质谱中呈现出独特的挑战。目前采用的方法是将蛋白质复合物溶解到膜类似物中,例如:去垢剂、纳米脂质盘、两性聚合物等,再将这些膜类似物通过碰撞激活去除。其中去垢剂是应用的最广泛的一种。然而由于碰撞激活的能量在应用中受到限制,该方法并不能在质量分析前很好地去除去垢剂。此外,在非变性质谱条件下,键的断裂也受到非共价相互作用强度的影响(例如蛋白质-蛋白质、蛋白质-去垢剂剂以及去垢剂胶束内的相互作用)。
基于光子的方法,如紫外光解离(UVPD)和红外多光子解离(IRMPD)已被证明有利于可溶性蛋白质及其复合物的Top-Down质谱分析。与此同时,基于光子的膜蛋白Top-Down模式的应用正在兴起。原理上,激光束路径中的离子被连续地驱动到振动激发态。因此,在基于光子的方法中,能量储蓄通常与前体离子的电荷状态和分子量无关。然而,电荷状态和分子量仍然会影响肽键解离需要的输入能量。先前报道的通过UVPD对79 kDa的五聚体的大电导机械敏感通道(MscL)Top-Down的断裂得到了令人印象深刻的54%的序列覆盖。然而,对于氨通道(AmtB)一个127 kDa的同源三聚体,通过碰撞激活和UVPD两种不同的方式破碎,仅实现了<20%的序列覆盖率。事实上,相对较低的序列覆盖率是由于大分子量以及三聚体中增加的非共价相互作用影响的结果。尽管这些工具能够在非变性状态下实现Top-Down质谱分析,但其在膜蛋白表征中的应用仍不广泛。这就要求建立一种能使低电荷密度的高分子量蛋白质稳定地产生蛋白质序列离子的方法,而膜蛋白嵌入异质膜或膜类似物则使这一问题更加复杂。虽然IRMPD之前被用于从去垢剂中释放膜蛋白,但使用IRMPD对非变性的膜蛋白进行测序的研究相对较少。
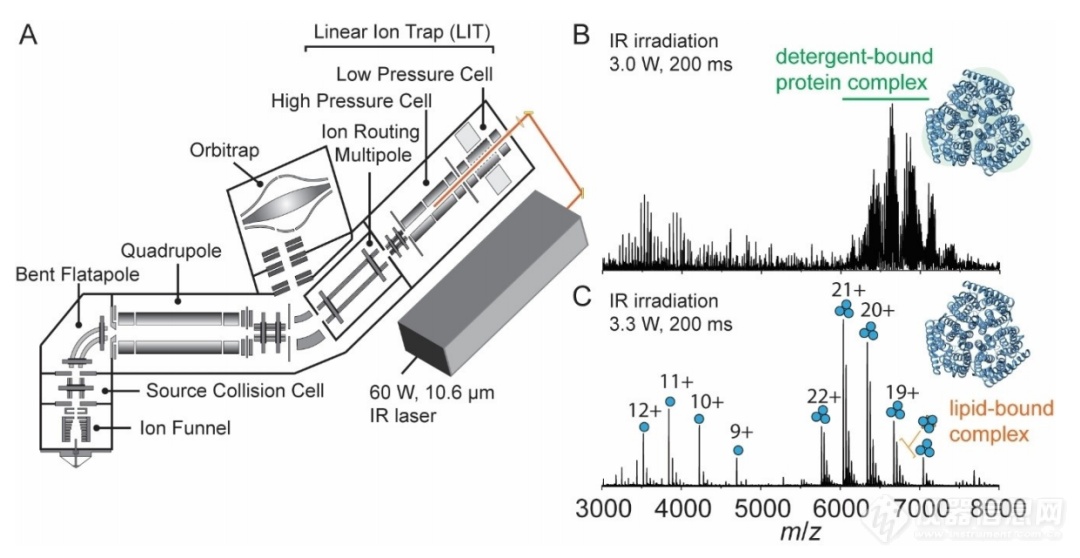
图1. (A)改进的Orbitrap Eclipse Tribrid的原理图,其中包括一个红外激光器直接进入四极线性离子阱(QLIT)的高压细胞。离子化的蛋白质胶束被转移到高压QLIT中,在那里整个离子群受到红外光子的照射,然后被转移到Orbitrap进行质量分析。通过调节激光输出功率(W)和照射时间(ms),可以使膜蛋白从去垢剂胶束中完全解放出来。(B)三聚氨通道(AmtB)在3.0 W输出功率和200ms辐照时间下的非变性质谱。(C)在3.3 W输出功率和200ms辐照时间下AmtB的非变性质谱。
因此,作者利用改进的Orbitrap Eclipse Tribrid质谱仪,与连续波远红外(IR) CO2激光器连接,使光束聚焦到双四极杆线性离子阱(QLIT)的高压池中(图1A)。红外激活可以有效地去除蛋白质复合物中的去垢剂胶束,随后通过IRMPD使得膜蛋白碎片化。在这种安排下,由纳米电喷雾电离产生的蛋白质复合物被转移到高压池中。在转移到Orbitrap进行检测或m/z分离和随后的碎片化之前,整个离子群将受到943cm-1红外光子的照射。利用红外的方法去除去垢剂胶束,红外激光有两个可调控参数:激光输出功率(高达60瓦)和照射时间(毫秒到秒)。因此,可以更好地控制从蛋白质胶束中释放膜蛋白,确保非变性复合物的保存,同时完全去除包裹复合物中的去垢剂。通过对激光输出功率和照射时间的优化,作者发现红外激活的参数是高度可调的,不同的激光功率和照射时间的组合也可以产生分辨率相当的谱图。其中例如在3.3 W下照射200 ms时,可以得到多个电荷态的三聚体峰(~6500 m/z),也可以观察到三聚体与脂质结合的峰,而且对于图谱中的单体也能观察到与脂质结合的峰(图1C)。作者还对不同的去垢剂产生分辨率较高的图谱所需要红外参数进行了评估,从而评价了这几种去垢剂得到高分辨率图谱的难易程度(图2)。
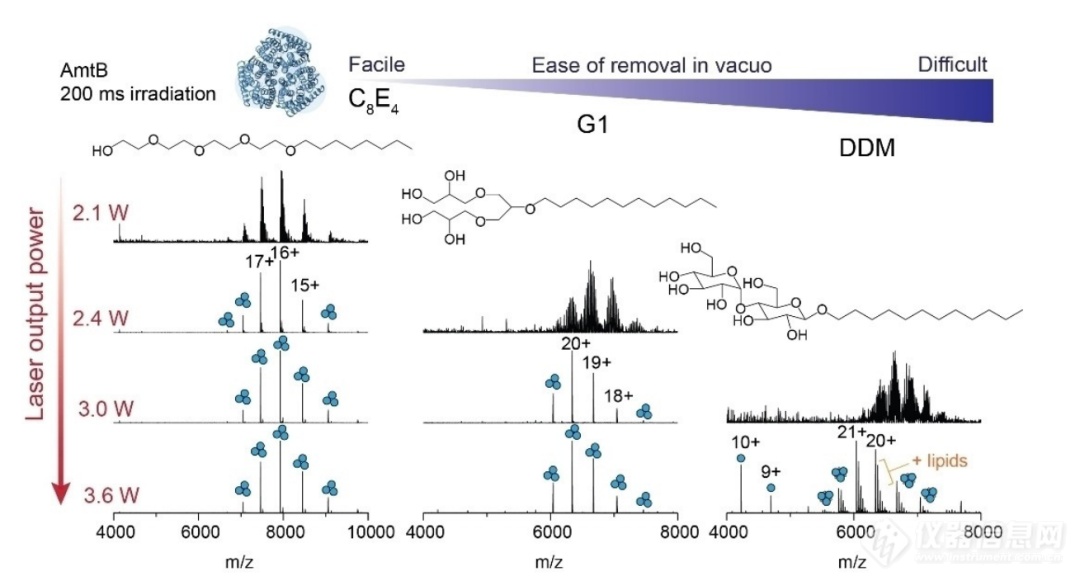
图2. 红外辐射去除膜蛋白离子中的去垢剂是高度可调的。增加激光输出功率对三种常用的MS兼容去垢剂(C8E4, G1和DDM) AmtB三聚体峰外观的影响。辐照时间固定为200 ms,激光输出功率分别为2.1、2.4、3.0和3.6 W。去垢剂在真空中按易去除的顺序显示,这是由完全释放膜蛋白复合物所需的激光输出功率决定的,从而在m/z光谱中产生良好分辨的电荷状态峰。为了探究IRMPD分离蛋白质和去垢剂胶束的机制,作者对三种不同的去垢剂:四聚乙二醇单辛醚(C8E4)、树突状低聚甘油(G1)和十二烷基-β-D-麦芽糖苷(DDM)的溶液相和气相红外光谱进行了表征,并利用密度泛函理论(DFT)计算得到了C8E4头部基团的红外谐波光谱,用来验证所得到的红外吸收光谱会受到振动耦合的影响,对于质子化的去垢剂离子,氢键和富氧去垢剂内的质子共享可以改变观察到的振动频率。结果表明C8E4胶束的溶液相吸收光谱包含一个与预期激光波数943cm-1重叠的显著带,这就解释了为何较低的激光能量可以将去垢剂胶束和蛋白质复合物分离。而在谐波光谱中在预期的激光波数处的确产生了峰,并推测该峰来自于O-H伸缩、C-C伸缩,C-H弯曲和C-O伸缩振动的耦合。而G1和DDM的最大吸收则偏离了943cm-1的预期波数,作者认为这是不同去垢剂氢键作用的结果。而蛋白质在真空中的红外吸收能力较弱,由此推测在IRMPD的过程中,去垢剂是主要的吸收对象。
所以仅需要较低的能量就可以使蛋白质从复合物中剥离而不至于破坏蛋白质的非共价作用。完整的蛋白质离子还支持串联质谱的实验,为了得到蛋白质的序列信息,作者分离了m/z在6674处(电荷态为+19)的AmtB三聚体蛋白,并将其置于高激光输出功率(9 W)下照射5 ms,在m/z 1750~4000之间产生密集的多电荷态离子片段,并得到了26%的序列覆盖,这优于之前基于碰撞激活的方法(<20%的序列覆盖率)。此外,在MS2的谱图中并没有观察到单体的峰,这说明共价键的断裂比蛋白质的展开和亚基的分离更快,这种效应也在之前的可溶性蛋白和膜蛋白研究中呈现。为了探究位点裂解的偏好,作者将片段离子丰度通过电荷态进行了归化一,发现了高频的断裂位点来自于经典的选择性断裂位点X|P和D|X,而剩余的断裂往往发生在A|G、F|G和V|G的位点,说明N端到甘氨酸有更高的断裂偏好。为了观察断裂的位置和蛋白质的二级结构之间的关系,作者沿着氨基酸序列构建了每个片段相对于原点位置的相对丰度图,多个电荷态的离子则通过归化一的方法进行求和。(图3)由此观察到了大多数的片段断裂出现在跨膜区域的α-螺旋处,其中8号α-螺旋的T|P和V|G,以及6号α-螺旋的L|G和F|G断裂产生了丰度最高的几个片段。此外,将这些片段的相对丰度映射到三聚体的结构上发现,片段来自于蛋白质的内部而非表面。分子动力学的研究表明了其中的机制,在高温下蛋白质的跨膜区域的α-螺旋保持了稳定的结构,而非跨膜区域的α-螺旋则失去了大部分的螺旋结构。先前的研究表明了α-螺旋外侧的质子化的氨基酸可以稳定α-螺旋的结构。由此,作者推测质子不光可以稳定蛋白质的螺旋结构,而且可以沿着蛋白质的骨架迁移来诱导电荷远程破碎。
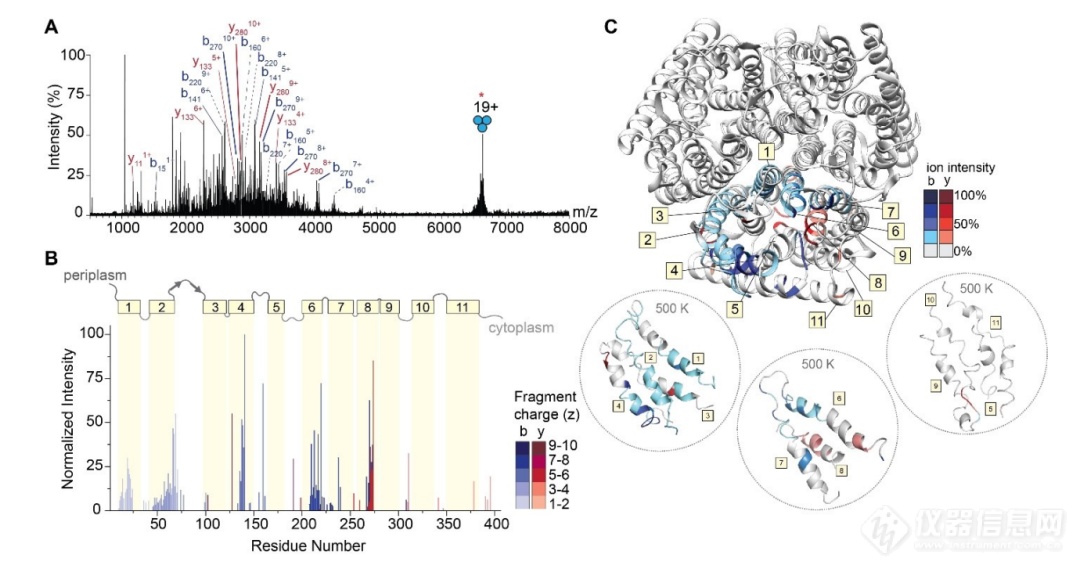
图3. 三聚体AmtB的IRMPD。(A)在m/z 6674处分离19+电荷态离子阱后,IRMPD后观察到的碎片离子MS2谱。多重带电碎片被高亮显示;来自相同地点的重复片段用虚线分组。为了清楚起见,许多指定的离子没有注释;(B)片段丰度相对于裂解原点(残基数)的条形图,其中丰度表示来自每个位点的片段归化一强度之和。条形图的颜色强度表示每个片段的加权平均电荷。将AmtB的拓扑域叠加在条形图上;α-螺旋跨膜区域用黄色方框表示,编号为1到11。跨膜区由质周环和细胞质环连接,用灰色线表示。(C)主干裂解位点覆盖在AmtB (PDB: 1U7G)的结构上。蓝色和红色阴影区域分别代表b型和y型离子。颜色强度对应于所分配片段的丰度。从气相分子动力学模拟中得到的高温(500 K)下的跨膜螺旋快照用虚线圈标出。
为了验证这一个推测,作者又对另外两种GPCR蛋白:β -1-肾上腺素能受体(β1AR)和腺苷A2A受体(A2AR)用IRMPD进行了MS2图谱的测定,结果也观察到了片段离子相似的二级结构定位,在跨膜结构区域有着高丰度的片段,但是在二硫键相连的螺旋中并没有观察到丰富的离子片段。并再次利用分子动力学模拟研究了两种GPCR的结构对断裂的影响。在500 K下的最终结构中显示,两种GPCR中都保留了α-螺旋特征,并与观察到的裂解位点密切相关。此外,还对这两种蛋白进行了HCD和IRMPD的比较分析。对于β1AR, IRMPD产生的片段离子平均分子量为8866 Da,高于HCD产生的5843 Da。IRMPD产生的片段离子也保留了更高的平均电荷(4.7 + vs 3.6+ z)。最终,IRMPD的碎片化导致β1AR的序列覆盖率更高,为28%,而HCD为17%。在A2AR中也观察到类似的趋势,IRMPD的覆盖率为19%,而HCD为9%。
总的来说,作者证明了可以在改进的Orbitrap Eclipse质谱仪的高压QLIT下,通过红外照射可以完全释放一系列去垢剂胶束中的膜蛋白。然后,通过增加激光输出功率,获得直接从膜蛋白及其复合物中释放的序列信息片段离子,证明红外光去除去垢剂是通用的和高度可控的,为保存和鉴定膜蛋白和配体之间脆弱的非共价相互作用构建了一个可靠的方法。而且还对片段离子的产生机制做了阐述,即质子可以通过沿蛋白质骨架迁移来稳定和诱导连续的肽键裂解。
撰稿:李孟效
编辑:李惠琳
文章引用:Infrared Multiphoton Dissociation Enables Top-Down Characterization of Membrane Protein Complexes and G ProteinCoupled Receptors
参考文献
Lutomski, C.A., El-Baba, T.J., Hinkle, J.D., et al. Infrared multiphoton dissociation enables top-down characterization of membrane protein complexes and g protein-coupled receptors[J]. Angewandte Chemie-International Edition,2023.
来源于:仪器信息网














热门评论
最新资讯
新闻专题
更多推荐
