推荐厂家
暂无
暂无

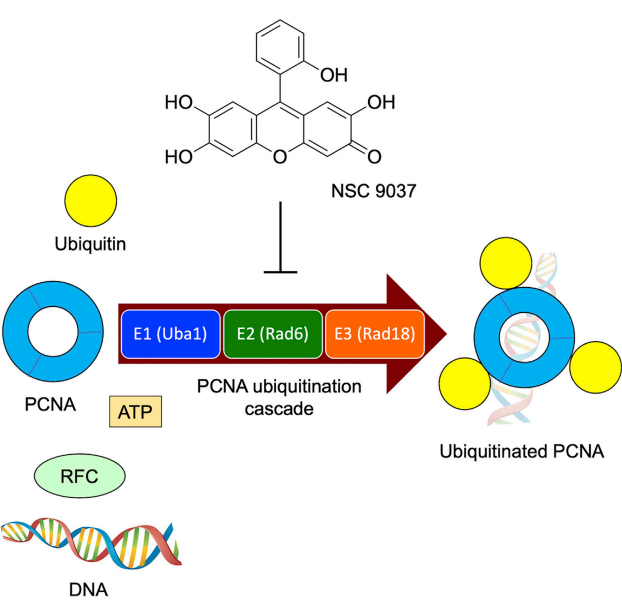



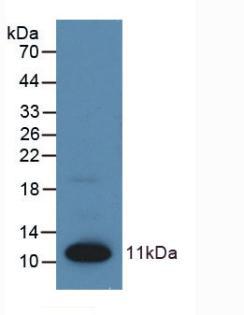
[font=宋体][font=宋体]泛素化是一种细胞内的蛋白质标记系统,蛋白质泛素化是指将小的蛋白质泛素共价地连接到其他蛋白质分子上的过程。泛素([/font][font=Calibri]ubiquitin[/font][font=宋体])是一种高度保守的蛋白质,其结构由[/font][font=Calibri]76[/font][font=宋体]个氨基酸残基组成。泛素连接到目标蛋白质上的过程,经历了泛素激活、泛素转移和靶蛋白接受三个主要步骤。[/font][/font][font=宋体] [/font][font=宋体]蛋白质泛素化具有多种特点,例如它是高度选择性的,不同蛋白质泛素化的位置和数量可以影响其功能;它是可逆的,通过去泛素化反应可以调控蛋白质的泛素化状态;它还是动态调控的,受到多种因素的调控,如细胞信号通路和环境刺激。[/font][b][font=宋体]泛素化蛋白大小:[/font][/b][font=宋体] [/font][font=宋体][font=宋体]蛋白泛素化是指将小蛋白颗粒泛素([/font][font=Calibri]Ubiquitin[/font][font=宋体])与其他蛋白质共价结合的修饰过程。 泛素化修饰通常会导致泛素共价连接在蛋白质的赖氨酸残基上形成多重泛素链。 这种蛋白质泛素化增加了蛋白质的分子量,因为每个泛素分子的质量大约为[/font][b][font=Calibri]8.5[/font][font=宋体]千达尔顿([/font][font=Calibri]kDa[/font][/b][font=宋体][b])[/b]。[/font][/font][font=宋体] [/font][font=宋体] [/font][b][font=宋体]泛素化蛋白质组学在许多领域有重要的应用,主要包括:[/font][/b][font=宋体] [/font][font=宋体]①疾病机制研究:泛素化是一种广泛存在于细胞中的蛋白质修饰方式,参与了细胞的生长、分化、修复和调控等多个生命活动。泛素化蛋白质组学的研究可以帮助我们了解泛素化修饰的生物学功能和调控机制,为疾病发生机制和治疗策略的研究提供重要线索。例如,在癌症、代谢综合征、神经退行性疾病等疾病中,则会出现异常泛素化。[/font][font=宋体]②药物研发:通过分析药物对泛素化蛋白质的影响,可以评估药物的效力和选择性,为药物研发提供指导。[/font][font=宋体]③临床诊断:泛素化蛋白质组学鉴定与定量分析技术可以揭示细胞调控的机制,通过分析泛素化蛋白质的组学数据,可以确定泛素化修饰在细胞信号转导、蛋白质降解和细胞周期调控等过程中的重要作用。此外,通过比较病态和正常样品中泛素化蛋白质的差异,可以鉴定与疾病发生发展相关的泛素化修饰靶点,并进一步理解疾病的分子机制。因此,这些技术也可用于临床诊断。[/font][font=宋体]④蛋白质降解调控:在癌症、神经退行性疾病和免疫相关疾病等病症中,蛋白质降解调控出现异常。而泛素化蛋白组在调控蛋白质降解中发挥重要作用。通过与泛素连接,目标蛋白质被送入蛋白酶体或蛋白酶体样体中进行降解。这个过程是细胞清除异常、老化或受损蛋白质的重要途径。[/font][font=宋体]⑤高通量技术应用:高通量泛素化蛋白质组学鉴定与定量分析技术的发展包括质谱鉴定和抗体鉴定两种方法。质谱鉴定技术利用质谱仪的高灵敏度和分辨率,能够鉴定泛素化修饰的蛋白质及其泛素化位点。抗体鉴定技术则通过特异性抗体的使用,可以富集和鉴定泛素化修饰的蛋白质。这些技术为全面了解泛素化在细胞中的作用机制和调控网络提供了可能。[/font][font=宋体]总的来说,泛素化蛋白质组学在多个领域都有重要的应用价值,推动了我们对生命过程的深入理解以及疾病治疗的创新发展。[/font][font=宋体] [/font][font=宋体]更多详情关于[url=https://cn.sinobiological.com/resource/protein-review][b]蛋白资源[/b][/url]详情可以参看:[/font][url=https://cn.sinobiological.com/resource/protein-review][u][font=宋体][color=#0000ff][font=Calibri]https://cn.sinobiological.com/resource/protein-review[/font][/color][/font][/u][/url][b][font=宋体] [/font][font=宋体][font=宋体]义翘神州:蛋白与抗体的专业引领者,欢迎通过百度搜索[/font][font=宋体]“义翘神州”与我们取得联系。[/font][/font][/b]
[size=15px][b][font=&][color=#0070c0]1[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]GA[/color][/font][font=宋体][color=#0070c0]诱导[/color][/font][font=&][color=#0070c0]KRAS[/color][/font][font=宋体][color=#0070c0]降解[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size][size=15px][font=宋体]作者首先通过筛选了一个自制的化合物库,用含有[/font][font=&] KRAS[sup]G12A[/sup][/font][font=宋体]突变的[/font][font=&]MM[/font][font=宋体]细胞系[/font][font=&]MM.1S[/font][font=宋体]和[/font][font=&]RPMI 8226[/font][font=宋体]鉴定可以降低[/font][font=&]KRAS[/font][font=宋体]水平的化合物,发现藤黄酸([/font][font=&]Gambogic acid[/font][font=宋体],[/font][font=&]GA[/font][font=宋体],)处理可降低两种细胞系中[/font][font=&]KRAS[/font][font=宋体]的水平,而其他化合物没有表现出相似或较弱的效果。进一步实验表明,[/font][font=&]GA[/font][font=宋体]可以以浓度和时间依赖性方式降低两种细胞系中[/font][font=&]KRAS[/font][font=宋体]的水平,以及[/font][font=&] KRAS[/font][font=宋体]的下游[/font][font=&]p-ERK[/font][font=宋体]([/font][font=&]MAPK[/font][font=宋体]通路)和[/font][font=&]p-AKT[/font][font=宋体]([/font][font=&]PI3K[/font][font=宋体]通路)。此外,[/font][font=&]q[url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url][/font][font=宋体]显示[/font][font=&]GA[/font][font=宋体]不会改变[/font][font=&]KRAS[/font][font=宋体]的转录水平,表明[/font][font=&]GA[/font][font=宋体]可能在转录后水平降低[/font][font=&]KRAS[/font][font=宋体]。[/font][font=&][/font][/size][size=15px][font=宋体]进一步使用蛋白酶体抑制剂[/font][font=&]MG132[/font][font=宋体]发现[/font][font=&]MG132 [/font][font=宋体]部分挽救了[/font][font=&]GA[/font][font=宋体]诱导的[/font][font=&]KRAS[/font][font=宋体]蛋白下调,表明[/font][font=&] KRAS [/font][font=宋体]蛋白降解与泛素[/font][font=&]-[/font][font=宋体]蛋白酶体途径有关,此外,我们采用放线菌酮([/font][font=&]CHX[/font][font=宋体])测定评估显示[/font][font=&]GA[/font][font=宋体]处理后,[/font][font=&]KRAS[/font][font=宋体]的半衰期显著缩短。这些数据表明[/font][font=&]GA[/font][font=宋体]可以诱导[/font][font=&]KRAS[/font][font=宋体]的蛋白酶体降解,进而损害[/font][font=&]MM[/font][font=宋体]细胞中的下游信号转导[/font][font=宋体] [size=15px][b]2、鉴定USP2作为GA的新靶标[/b][/size][size=15px]为了确定GA诱导KRAS降解的靶标,作者合成了生物素标记的GA,发现它保留了在MM细胞中诱导KRAS降解的能力,进一步利用该探针开展Pulldown+MS,鉴定到的互作蛋白中,HSP90和USP2这两种与蛋白质降解相关。HSP90是一个已知的GA靶点,作为伴侣蛋白,它可以调节各种蛋白的稳定性,然而HSP90抑制剂STA9090处理不能降低MM.1S细胞中KRAS的蛋白水平,表明KRAS不是 HSP90的底物。因此,作者更加关注USP2,一种新型的GA互作蛋白。通过Pulldown+WB、免疫荧光共定位、竞争实验、CETSA、DARTS实验共同证实了USP2是GA的直接相互作用蛋白。此外,GA在体外相对特异性地抑制USP2的去泛素化活性( [/size][size=15px][b]3、半胱氨酸284对于GA与USP2的共价结合至关重要[/b][/size][size=15px]然后,作者研究了GA和USP2之间的相互作用模式,发现碘乙酸(IAA,一种半胱氨酸烷化剂)与USP2的预孵育完全抑制了GA与USP2的结合,表明GA可能与USP2的半胱氨酸共价结合,这与之前的报道一致,即GA可以通过半胱氨酸残基与其靶标共价结合。接着,通过USP2蛋白与GA一起孵育后质谱鉴定,发现Cys284残基被GA共价修饰。此外,通过构建USP2的两个突变体(C276S、C284S),发现GA 与 USP2(C276S)结合,而不是USP2(C284S),表明GA可以与USP2的Cys284残基特异性形成共价键。结合动力学显示GA 以剂量和时间依赖性方式与 USP2 共价结合,分子对接显示P565和 R289对于形成GA与USP2结合的口袋至关重要。同样通过蛋白点突变实验发现P565 和 R289对GA-USP2相互作用至关重要 [/size][size=15px][b]4、USP2调节KRAS的稳定性[/b][/size][size=15px]前面发现GA降低MM细胞中KRAS蛋白水平并抑制USP2活性,表明USP2可能调节KRAS的稳定性。作者利用CRISPR/Cas9敲除MM.1S和RPMI 8226细胞系中的USP2,发现敲除USP2导致KRAS蛋白水平降低,mRNA水平不变, KRAS的下游效应子p-ERK水平也显著降低。此外,敲除诱导的KRAS下调可被两种细胞系中的蛋白酶体抑制剂MG132挽救。结果表明USP2可以增强KRAS的蛋白质稳定性。进一步研究发现USP2与KRAS互作并使KRAS去泛素化,表明KRAS是USP2的底物 [/size][size=15px][b]5、USP2敲除抑制MM细胞的增殖[/b][/size][size=15px]鉴于KRAS在MM细胞增殖和存活中的关键作用,作者假设USP2敲除可能会使KRAS不稳定,从而导致MM细胞增殖抑制。结果显示USP2敲除显著抑制MM.1S和RPMI 8226细胞系的增殖,诱导这两种MM细胞系凋亡。此外,MM.1S 细胞的异种移植 MM 模型发现,USP2敲除显著抑制肿瘤生长,免疫组化染色也显示USP2敲除组的KRAS水平降低,这些数据共同表明USP2在MM细胞的增殖和存活中起着关键作用 [/size][size=15px][b]6、GA通过靶向USP2和破坏KRAS的稳定性来诱导MM细胞凋亡[/b][/size][size=15px]据报道,GA通过抑制PI3K/Akt/mTOR、[i]NF-κ[/i]B 和其他信号通路诱导MM细胞凋亡。作者发现GA处理可以以剂量依赖性方式降低MM.1S和RPMI 8226细胞的活力并促进细胞凋亡。此外,过表达USP2的细胞对GA诱导的活力降低和KRAS降解表现出部分抗性,且GA处理可以提高 KRAS的泛素化水平。KRAS[sup]G12C[/sup]的过表达可以部分消除USP2敲除诱导的细胞生长抑制。这些数据表明,GA在 MM 细胞中诱导的细胞毒性至少部分归因于其靶向USP2,这降低了KRAS的稳定性。[/size][size=15px]最后,作者探讨了USP2在MM中的临床意义。通过GSE13591数据集发现MM患者骨髓样本中的USP2 mRNA水平明显更高。此外,作者发现原发性骨髓瘤细胞和骨髓瘤细胞系中USP2的蛋白水平高于正常外周血单核细胞(PBMC)和正常骨髓活检,且高USP2水平的MM患者的总生存期降低,这些结果共同表明USP2表达增加与MM中较差的结局之间存在很强的相关性,表明USP2可能是MM治疗的有前途的靶点。[/size][/font][font=&][/font][/size]
[size=15px][font=宋体][color=black]冬凌草乙素[i][/i]([/color][/font][font=&][color=black]Ponicidin[/color][/font][font=宋体][color=black])是从中药冬凌草([/color][/font][i][font=&][color=black]Rabdosia rubescens[/color][/font][/i][font=宋体][color=black])中提取的二萜类化合物,具有免疫调节、抗炎、抗病毒和抗癌等多种活性。尽管冬凌草乙素对多种恶性肿瘤有疗效,但其与肝细胞癌([/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black])相关的确切功能和作用机制仍然未知。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=宋体][color=black]冬凌草乙素体外显著抑制肝癌细胞增殖和迁移,体内抑制肿瘤生长并促进肿瘤细胞凋亡。[/color][/font][font=宋体][color=red]机制上,冬凌草乙素靶向[/color][/font][font=&][color=red]Keap1[/color][/font][font=宋体][color=red]([/color][/font][font=&][color=red]E3[/color][/font][font=宋体][color=red]泛素连接酶)并促进[/color][/font][font=&][color=red]Keap1-PGAM5[/color][/font][font=宋体][color=red]复合物形成,介导[/color][/font][font=&][color=red]PGAM5[/color][/font][font=宋体][color=red]的泛素化降解。此外,冬凌草乙素通过[/color][/font][font=&][color=red]PGAM5[/color][/font][font=宋体][color=red]激活半胱氨酸依赖性线粒体通路,导致线粒体损伤和[/color][/font][font=&][color=red]ROS[/color][/font][font=宋体][color=red]产生,从而促进肝癌细胞线粒体凋亡。[/color][/font][font=&][color=black][/color][/font][/size] [align=center] [/align] [size=15px][b][font=&][color=#0070c0]1[/color][/font][font=宋体][color=#0070c0]、冬凌草乙素抑制[/color][/font][font=&][color=#0070c0]HCC[/color][/font][font=宋体][color=#0070c0]细胞的增殖和迁移[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [align=center] [/align] [size=15px][font=宋体][color=black]作者首先通过体外实验发现能以剂量依赖性方式有效抑制[/color][/font][font=&][color=black]HepG2[/color][/font][font=宋体][color=black]细胞[i][/i]的增殖和迁移。为了确定冬凌草乙素的靶标,作者合成生物素标记的冬凌草乙素([/color][/font][font=&][color=black]Bio-Ponicidin[/color][/font][/size][font=宋体])开展[/font][font=宋体]Pulldown[/font][font=宋体]实验,通过质谱鉴定[/font][font=宋体]Keap1[/font][font=宋体]蛋白([/font][font=宋体]Kelch-like ECH-associated protein 1[/font][font=宋体],[/font][font=宋体]Keap1[/font][font=宋体],[/font][font=宋体]E3[/font][font=宋体]泛素连接酶的底物识别亚单位)为冬凌草乙素的可能靶标。[/font] [align=center] [/align] [size=15px][b][font=&][color=#0070c0]2[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]PGAM5[/color][/font][font=宋体][color=#0070c0]在[/color][/font][font=&][color=#0070c0]HCC[/color][/font][font=宋体][color=#0070c0]组织样本中上调[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]接着,作者利用[/color][/font][font=&][color=black]TCGA[/color][/font][font=宋体][color=black]数据库发现[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]高表达与[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]患者较低的生存率有关,并利用[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]组织芯片发现肝癌组织中[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]的表达高于癌旁组织,结果表明[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]在[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]发病机制中具有潜在作用。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]蛋白是一种重要的调节蛋白,可以通过与其他蛋白质相互作用来调节细胞内信号通路,于是作者通过文献检索发现[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]是一种与[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]互作的重要蛋白质,且前面的[/color][/font][font=&][color=black]Pulldown[/color][/font][font=宋体][color=black]实验也显示[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]被拉下。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=&][color=black]TCGA[/color][/font][font=宋体][color=black]数据库分析显示[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]高表达与[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]存活率较低相关,组织芯片显示[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]组织中的[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]表达高于癌旁组织,且与较高的病理分级相关,结果表明[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]同样在[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]发病机制中具有潜在作用。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=宋体][color=black]进一步作者通过人类蛋白质组微阵列[i][/i]检测冬凌草乙素的直接靶蛋白,发现冬凌草乙素与[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]直接结合而不与[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]蛋白结合,结果表明冬凌草乙素可能直接与[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]结合并影响[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black],从而在[/color][/font][font=&][color=black]HCC[/color][/font][font=宋体][color=black]中发挥药理作用。[/color][/font][font=&][color=black][/color][/font][/size] [align=center] [/align] [size=15px][b][font=&][color=#0070c0]3[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]Keap1[/color][/font][font=宋体][color=#0070c0]和[/color][/font][font=&][color=#0070c0]PGAM5[/color][/font][font=宋体][color=#0070c0]相互作用的结构基础[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]可以与[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]结合,然而,它们结合的结构基础尚不清楚。为了观察[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]与[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]结合过程的动态变化,作者通过分子动力学模拟发现[/color][/font][font=&][color=black]Keap1-PGAM5[/color][/font][font=宋体][color=black]复合物的结构总体上保持稳定,且[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]上的[/color][/font][font=&][color=black]Val78[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]Glu79[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]Ser80[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]Glu83[/color][/font][font=宋体][color=black]氨基酸与[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]的[/color][/font][font=&][color=black]Kelch[/color][/font][font=宋体][color=black]结构域相互作用。采用[/color][/font][font=&][color=black]AlphaFold3[/color][/font][font=宋体][color=black]算法来预测[/color][/font][font=&][color=black] Keap1-PGAM5 [/color][/font][font=宋体][color=black]的相互作用,发现复合物的总体预测折叠与真实结构相似。[/color][/font][font=&][color=black][/color][/font][/size] [align=center] [/align] [size=15px][b][font=&][color=#0070c0]4[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=&][color=#0070c0]Keap1-PGAM5[/color][/font][font=宋体][color=#0070c0]配合物中晶体整体结构及相互作用的洞察分析[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]为了更好地理解[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]和[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]相互作用的分子机制,作者进行了结构生物学实验。通过晶体学实验获得了[/color][/font][font=&][color=black]Keap1-PGAM5[/color][/font][font=宋体][color=black]配合物的结构,分析得到两者的结合模式和结合位点,并通过蛋白点突变后的[/color][/font][font=&][color=black]ITC[/color][/font][font=宋体][color=black]实验发现[/color][/font][font=&][color=black]Glu79[/color][/font][font=宋体][color=black]是[/color][/font][font=&][color=black]Kelch[/color][/font][font=宋体][color=black]与[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]结合的关键残基。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=宋体][color=black]进一步作者通过[/color][/font][font=&][color=black]SPR[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]CETSA[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]Co-IP[/color][/font][font=宋体][color=black]、[/color][/font][font=&][color=black]EMSA[i][/i][/color][/font][font=宋体][color=black]等实验验证冬凌草乙素和[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]的[/color][/font][font=&][color=black]Kelch[/color][/font][font=宋体][color=black]结构域结合,而不能和[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]的[/color][/font][font=&][color=black]?54-PGAM5[/color][/font][font=宋体][color=black]([/color][/font][font=&][color=black]54-289[/color][/font][font=宋体][color=black]号氨基酸)突变蛋白结合。[/color][/font][font=&][color=black][/color][/font][/size] [size=15px][font=宋体][color=black]考虑到[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]是一种[/color][/font][font=&][color=black]E3[/color][/font][font=宋体][color=black]连接酶,促进蛋白质的泛素化和降解。作者发现冬凌草乙素可以增加[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]的泛素化,增加[/color][/font][font=&][color=black]Keap1-PGAM5[/color][/font][font=宋体][color=black]蛋白共定位,表明冬凌草乙素可以与[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]结合,从而促进[/color][/font][font=&][color=black]Keap1[/color][/font][font=宋体][color=black]与[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]的互作,促进[/color][/font][font=&][color=black]PGAM5[/color][/font][font=宋体][color=black]的泛素化。 [/color][/font][/size] [size=15px][b][font=&][color=#0070c0]5[/color][/font][font=宋体][color=#0070c0]、[/color][/font][font=宋体][color=#0070c0]冬凌草乙素影响[/color][/font][font=&][color=#0070c0]Keap1[/color][/font][font=宋体][color=#0070c0]的[/color][/font][font=&][color=#0070c0]Kelch[/color][/font][font=宋体][color=#0070c0]结构域的变构来稳定[/color][/font][font=&][color=#0070c0]Keap1[/color][/font][font=宋体][color=#0070c0]与[/color][/font][font=&][color=#0070c0]PGAM5[/color][/font][font=宋体][color=#0070c0]结合[/color][/font][font=&][color=#0070c0][/color][/font][/b][/size] [size=15px][font=宋体][color=black]进一步通过分子对接模拟发现冬凌草乙素可以与[/color][/font][font=&][color=black]Keap1[/color][/font]

 400-860-5168转3826
400-860-5168转3826
 留言咨询
留言咨询
 400-860-5168转3826
留言咨询
400-860-5168转3826
留言咨询
 400-860-5168转3541
留言咨询
400-860-5168转3541
留言咨询









 我要推广仪器
我要推广仪器
 下载APP
下载APP