仪器信息网APP
选仪器、听讲座、看资讯
立即体验
APP内打开
回版面
评论
收藏
点赞
拍砖
举报
取消
发布
当前位置:
仪器社区
>
生命科学仪器
>
生命科学仪器综合讨论
>
帖子详情
机械力作用下DNA结构变化的实时可视化
QUANTUM量子科学
2018/08/02
私聊
生命科学仪器综合讨论
研究DNA过度拉伸时的结构变化
单分子力谱(SMFS)仪器被广泛用于研究DNA过度拉伸时的结构变化。然而,很多仪器仅仅能够给出大体的信息,然而结构变化的作用机制仍然不明。将SMFS与可视化相结合可以解决这个问题。我们将讨论LUMICKS的C-Trap技术如何将高分辨率的光镊与荧光共聚焦显微结合来检测机械力作用下双链DNA至单链DNA的转变过程。
之前的研究表明,裸露的双链DNA在大约65 pN的力下发生转变。在施加恒定力的转变过程中,DNA的伸直长度增加了70%。接下来DNA的B型至S型转变中,包括若干因终端或缺口被剥开或因融解泡形成而融解的区域。由于DNA的这种行为在65 pN力作用下可以再现,因而可以测量不同的连接配体或不同的缓冲液产生的力导致的过度拉伸时的变化,揭示DNA与连接配体的相互作用如何影响DNA的结构与稳定性。
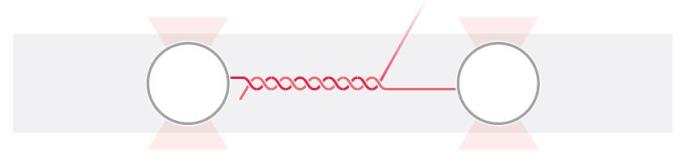
1 DNA过度拉伸的图示。DNA连接两个被光阱捕获而分开的微球,诱导了两端双链解旋分离。
我们使用高分辨率光镊操控DNA分子引发转变,同时检测力(亚pN级)和距离(亚nm级)随时间发生的变化。我们使用荧光标记来区分已经融解的单链DNA和双链DNA,并用高精度的(误差在15 nm以下)多通道单光子检测来确定融解的位置。使用层流微流控和自动装载功能,可用两个被光阱捕获的微球体来拴住双链DNA(图1)。接下来这个整体被输送至含有浓度为2nM的特异性结合双链DNA的Sytox橙和3 nM的荧光标记的特异性结合单链DNA的复制蛋白A(RPA)的通道中。
2 双链DNA转变至单链DNA的不同阶段的双色二维荧光共聚焦图像。绿色表示特异结合双链DNA的Sytox橙的荧光,蓝色表示特异结合单链DNA的RPA的荧光。
第一个实验中拍摄了不同声力下的荧光共聚焦图像以演示系统的双通道检测功能。首先我们用30 pN的力使双链DNA可见(图2a),碱基对的距离使得Sytox分子可以充分结合并完全包被双链DNA。第二个实验中,施加逐渐上升的声力直到过度拉伸,双链DNA从两端开始剥离。实验中观察到了一个非常明显的与RPA结合到单链DNA的区域(图2,蓝色区域)。除了两端以外的RPA结合的DNA区域显示了从DNA缺口处开始的剥离现象的存在。接下来用C-Trap微流控系统验证了DNA融解区域的存在,并对DNA施以缓慢的垂直流动缓冲液来延长剥下的单链DNA(图2C)。最后,双链DNA被声力完全融解,形成完全被RPA包被的单链DNA(图2D)。
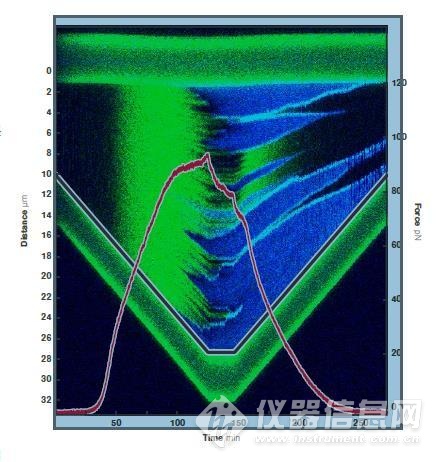
3 单个双链DNA分子以140 nm/s的速度拉伸与收缩的双色荧光波动曲线。将距离(灰色)与力(红色)的数据与荧光图像进行叠加得到数据的真实相关。
为使荧光显微结果与定量力谱数据相符,将单个双链DNA分子以140 nm/s的恒定速度进行拉伸并记录力、距离和荧光(图3)。
灰色的线显示了DNA两端距离随时间的变化,红色的线显示了施加的力随时间的变化。将所有的数据叠加,获得了每一条荧光显微的线的力和距离的值。因此,可以得出Sytox橙最低在25 pN的力下开始结合双链DNA。还有,和裸露的双链DNA不同,直到力达到90 pN才观察到了过度拉伸导致的变化。这样的现象可能是因为Sytox橙结合双链DNA稳定了结构,使其更加难于融解。当过度拉伸导致变化时产生了蓝色荧光,显示出双链DNA到单链DNA的结构变化。而且又一次观察到DNA的剥离不仅从两侧而且从缺口处开始的现象。
双链DNA开始融解,RPA开始结合单链DNA时,力的大小开始下降,表示包被了RPA的融解DNA已经稳定,不能再次退火形成双链DNA。最后,力的大小降至20 pN,Sytox橙分离,RPA依然保持结合。在理解了小分子配体存在时的DNA的结构变化和如何通过分子水平的机械操纵来研究这些相互作用可以引导生物学和生物物理学产生突破性的发现。
使用C-Trap光镊-荧光技术可以实时观察并检测结构变化。单分子的张力和拉伸的数据对研究生物分子和生物聚合物有重大意义。
相关话题
1
五颜六色的采血管
2
人用肝素化采血管的质量控制
3
生气与疝气有关吗
4
硅胶假体-乳房植入物的安全性
5
《科学》癌症药物治疗的效果取决于营养微环境:忌口很重要!
近期热榜
赛默飞实验室产品焕新计划有奖调研!
【七月征文】不一YOUNG实验“猿”
报名开启:ICS2024第十三届光谱网络会议!
推荐收藏!盘点中药材及饮片检测解决方案
热门活动
甄选国优仪器,助推设备更新
采购咨询618活动:1000元奖励等你拿
猜你喜欢
最新推荐
热门推荐
更多推荐
【第三届原创参赛】血液中总胆固醇(CHOL)测定COD-PAP方法验证报告
2010/09/11
毛细管气相色谱法:高分辨率,柱效高,开管型→挥发油
分享
2012/02/22
Cell:鉴定出全基因表达分析标准方法存在重大缺陷
分享
2012/11/03
哪个液氮罐品牌比较便宜
已应助
2024/01/17
自增压液氮罐“最新市场价格”的解密与应用
原创
2023/12/20
【分享】生物实验室常用技术参数资料
2008/11/24
Nature:清华大学宫鹏教授认为传统文化阻碍中国科研进步
讨论
2012/02/13
3万吨糖蜜酒精生产DCS控制系统
分享
2012/04/14
【我们不一YOUNG】结缘仪器,一起结缘,与仪器信息网一起成长
原创
2024/07/01
心血管检测仪打开后出现ch375dll.dll电脑丢失怎么办
已应助
2024/07/01
出售贝克曼DXC800生化分析系统
原创
2024/06/21
编织袋拉力机
原创
2024/06/14
求助贴: AKTA锯齿型洗脱曲线的解决办法
求助
2024/06/02
5-6折安捷伦耗材优惠价出售
分享
2024/05/30
液氮补给罐静态蒸发率范围解析
原创
2024/05/21
杂交瘤技术的应用及局限性
资料
2024/05/20
实验室检测记录能否使用Excel保存
求助
2024/06/28
仪器点火失败
求助
2024/06/28
求助,非甲烷总烃测定中,除烃空气0.4mg/m3,怎么计算的呢?是按总烃计算公式吗?
求助
2024/06/29
检测报告一定要有判定结果吗?
求助
2024/07/01
求助安捷伦GC7890B,EPC无响应解决办法
求助
2024/07/01
HJ1332便携非甲实际应用问题
讨论
2024/07/01
衍射斑点标定求助
求助
2024/06/30
气相FID重现性不好原因
求助
2024/07/01
安捷伦6470液质使用问题
讨论
2018/08/02
我的纺织品实验室配制和标定标准溶液分八步走
第十一届原创
2018/08/02
仪器自稀释与人工稀释之比较测试
第十一届原创
2018/08/02
应届毕业生‘遇见’面试官老司机的‘陷阱’怎么破!
第十一届原创
2018/08/02
使用光镊在单分子水平检测蛋白折叠、去折叠和构象动力学
分享
2018/08/02
清洗分流平板
第十一届原创
2018/08/02
如何提高气溶胶输送效率?
讨论
2018/08/02
为医药研发打造利器!月旭推出医药行业方法开发工具包
分享
2018/08/02
品牌合作伙伴
岛津
日立科学仪器
珀金埃尔默仪器(上海)有限公司(PerkinElmer)
日本电子株式会社
丹纳赫
安捷伦
赛默飞世尔科技
普析通用
欧波同
天美
天瑞仪器
德国耶拿
海能技术
马尔文帕纳科
磐诺科技
上海仪电科仪
梅特勒托利多
聚光科技
莱伯泰科
盛瀚
多宁生物
丹东百特
科哲
卓立汉光
屹尧科技
华谱科仪
宝德仪器
优莱博
HORIBA
布鲁克核磁
举报帖子
执行举报
点赞用户
好友列表
加载中...
正在为您切换请稍后...