

冰结构蛋白的性质、应用及限量标准
摘 要:冰结构蛋白(ISP)因其能影响冰晶生长规律的特性,被广泛用于食品添加领域中。 本文主要探讨总结冰结构蛋白的分类、抗冻机制、应用、限量、标准及检测方法。
关键词:冰结构蛋白;食品添加剂;抗冻机制
1、
2、 冰结构蛋白的性质和应用
1. 冰结构蛋白的基本性质
冰结构蛋白(Ice Structuring proteins,ISP),又称抗冻蛋白或热滞蛋白,是一种能防止冰结晶,控制其生长,改变其生长规律、速度、冰芯和冰晶在冻结或自然生长过程中的形状和大小,抑制重结晶的天然蛋白质。它们可以非依数性(跟其浓度无关)降低溶液的凝固点,而对熔点的影响很小,从而导致溶液的凝固点和熔点之间的差异,此差值称为热滞活度(thermal hysteresis activity,THA)。ISP存在于动物(昆虫、鱼类等)、植物(胡萝卜、冬小麦草、桃树等)、细菌和真菌中,保护生物有机体免受低温冻害。
2. 冰结构蛋白的分类[1]
从20世纪70年代首次从鱼类的血液中提取出冰结构蛋白以来,经过几十年的研究发展,人们相继从植物,昆虫,细菌,真菌中发现了具有同样功能的蛋白质,它们在组成和结构上各不相同,并在基因序列上也没有同源性,从而造就了冰结构蛋白的多样性和复杂性。鱼类中冰结构蛋白主要分为6大类,分别是ISGP(Ice Structuring Glycoprotein)、ISPⅠ、ISPⅡ、ISPⅢ、ISPⅣ、Hyperactive-ISP。目前研究最多的昆虫ISP基本都是从黄粉虫(Tenebrio Molitor)、毛虫(Dendroides Canadensis)、枞色卷蛾(Choristoneura Fumiferana)、美洲脊胸长椿(Oncopeitus Fasciatus)等4种昆虫中分离纯化的。昆虫ISP是目前已知热滞活性最高的ISP,可达5℃~10℃。植物中冰结构蛋白的研究比较晚,但是其来源广泛易得,如胡萝卜,女真叶,冬小麦等都可提取冰结构蛋白,并且植物性的冰结构蛋白在应用中也容易为人们所接受,因此对植物性冰结构蛋白的研究越来越多,如江南大学的黄卫宁等主要就是研究植物性冰结构蛋白的提取以及在冷冻面食中的应用。植物的冰结构蛋白的抗冻活性不高,热滞活性略低于极地鱼类的冰结构蛋白活性。
从细菌中分离提纯得到的纯冰结构蛋白研究很少。从南极洲的细菌中有6种细菌可分离出ISP,从菌株Moraxella分离出的ISP为脂蛋白,是第一次报道的冰结构脂蛋白;E.coli的ISP分子量大约为7.4ku,称为CS74,含大量的亲水基团;研究者还在加拿大北极地区的一种根瘤菌(Pseudomonasputida GR 12-2)中发现了一种冰结构蛋白。真菌中冰结构蛋白的研究也非常少,有报道称冬菇(Flammulinavelupites)和牡蛎菇(Pleurotusostreatus)中存在冰结构蛋白。
种类 | 来源 | 分子量(kD) | 特点 |
ISGP | 鱼类 | 2.6-33.7 | 糖基是主要作用基团 |
ISPI | 鱼类 | 3.3-4.5 | 只能在细胞中作用 |
ISPII | 鱼类 | 11-24 | 可能需要Ca2+稳定 |
ISPIII | 鱼类 | 6.5-14 | |
ISPIV | 鱼类 | ~12.3 | 富含谷氨酸 |
Hyperactive-ISP | 鱼类 | 活性远强于ISPI | |
黄粉虫ISP | 昆虫 | 8.4 | 热滞活性远高于鱼类ISP(10-100倍) |
毛虫ISP | 昆虫 | 8.7 | 耐高温(>100℃) |
枞色卷蛾ISP | 昆虫 | 9.0 | 抗冻活性高 |
胡萝卜ISP | 植物 | 36.8 | |
黑麦草ISP | 植物 | 13 | 耐高温(>100℃) |
沙东青ISP | 植物 | 形式多样、协同作用 | |
PCA60 | 植物 | 50 | 强冰晶修饰能力 |
细菌ISP | 细菌 | 研究很少 | |
真菌ISP | 真菌 | 研究很少 |
3. 冰结构蛋白的抗冻机制
冰结构蛋白主要通过热滞效应、重结晶抑制效应来发挥它的抗冻活性。热滞效应是指冰结构蛋白可以非依数性的降低溶液的冰点,而对其熔点基本没有影响,从而使冰点与熔点之间出现一差值,这个差值的大小就反映了冰结构蛋白的热滞活性,差值越大活性越高。重结晶抑制效应是指冰结构蛋白可以抑制冰晶重结晶,而重结晶是指由于温度波动,已形成的冰晶颗粒进行重新分配,大的冰晶越来越大,小的越小。
冰结构蛋白的抗冻机制主要分为吸附抑制模型、结构互补模型(受体-配体模型)、晶格匹配模型、偶极子-偶极子模型、晶格占有模型。冰结构蛋白种类繁多、同源性小且分子量差异大,故实际抗冻机制是复杂且多样的。
吸附抑制模型[2]指出,冰结构蛋白可以吸附到特定的冰晶c轴表面,通过Kelvin效应抑制冰晶在α方向的生长,从而减缓冰晶生长速率,以修饰冰晶形态。吸附抑制模型尚不能完整解释冰结构蛋白抑制冰晶生长的机理,即冰结构蛋白和冰晶间不仅有相互作用关系。结构互补模型指出[3],冰结构蛋白和冰晶表面通过不可逆的疏水作用和范德华力结合,从而法绘抗冻活性。
4. 冰结构蛋白的安全性研究[4][5]
(1) 氨基酸序列分析
BLAST(局部序列比对基本检索工具)和PIR-NREF(蛋白质信息库)测序结果表明,冰结构蛋白与抑制过敏原无同源性。
(2) IgE和组织胺含量测定
人体中的免疫球蛋白E(IgE)的含量一般很低,且嗜碱性粒细胞一般不会释放组织胺。而过敏性物质可以导致IgE和组织胺的含量增加。因此,通过测定IgE和组织胺的浓度变化,可以检验待测物质是否是过敏原。放射免疫吸附法(RAST)表明添加冰结构蛋白后不会造成IgE含量的上升和组织胺的释放,因此冰结构蛋白不会引起明显的过敏反应。
5. 冰结构蛋白在食品中的应用
(1) 冰结构蛋白在冰淇淋生产中的应用
所有冰淇淋行业仍然面临如何减少冷冻过程中的冰晶的生长和重结晶的问题,尤其对于中低端冰淇淋。冰结构蛋白的研究让人们看到希望。ISP可以结合冰晶表面,防止冰晶形成,控制其生长,改变生长规律,生长速度和形状和大小,并能抑制重结晶,从而避免产生大冰晶而影响口感,并能提高其稳定性。2006年,我国首次在国家食品安全标准GB2760中表明,可以冰结构蛋白作为新型食品添加剂添加到冷冻食品中。国际上,目前只有联合利华将冰结构蛋白应用到冰淇淋中。
(2) 冰结构蛋白在冷冻面团生产中的应用[6]
冷冻面团技术是一门新的烘焙食品加工技术,面团经过长时间的冷藏,面团的品质会下降,如醒发时间延长,比容减小等。添加0.5%ISP的面团可以有效的缩短面团的醒发时间,增大面包比容而减小面包硬度。扫描电镜结果表明,添加ISP的面团经长时间冻藏后,仍能保持其面筋网络结构。
(3) 冰结构蛋白在肉制品生产中的应用[7]
肉制品经过反复冷冻,由于冰晶不断集聚变大,会对肉制品的组织产生破坏,从而使汁液外漏,营养流失。而加了ISP的肉制品或者在宰杀前24h将0.01mg/mL的ISP注射入待宰羔羊体内,均可有效地减少渗水和冰晶的形成,从而保持其原有的组织结构,减少营养流失。这可能是因为ISP使部分融化的冰晶缓慢地渗入到细胞内,在蛋白质颗粒周围重新形成水化层,使汁液流失减少,保持解冻食品的营养成分和原有风味。
(4) 冰结构蛋白在农产品生产中的应用
随着各种ISP的基因测序的进行,研究者可通过基因工程技术将ISP的基因导入植物体内,并在体内表达,使植物获得抗冻活性,选育出转基因的抗冻植物,从而扩大植物的种植范围。
3、 冰结构蛋白的限量、检测和标准
项目 | 标准 | 检测 |
感官 | 浅棕色液体 | - |
ISP含量(g/L) | ≥5 | HPLC或MS |
灰分(%) | ≤2 | GB5009.4 |
pH | 2.5-3.5 | pH计 |
铅(mg/L) | ≤2 | GB5009.75 |
菌落总数(CFU/mL) | ≤3000 | GB4789.2 |
大肠菌群(CFU/mL) | ≤10 | GB4789.3 |
霉菌和酵母菌(CFU/mL) | ≤100 | GB4789.15 |
沙门氏菌(25mL) | 不得检出 | GB4789.4 |
单核细胞增生李斯特氏菌(25mL) | 不得检出 | GB4789.30 |
蜡样芽孢杆菌(CFU/mL) | ≤100 | GB4789.14 |
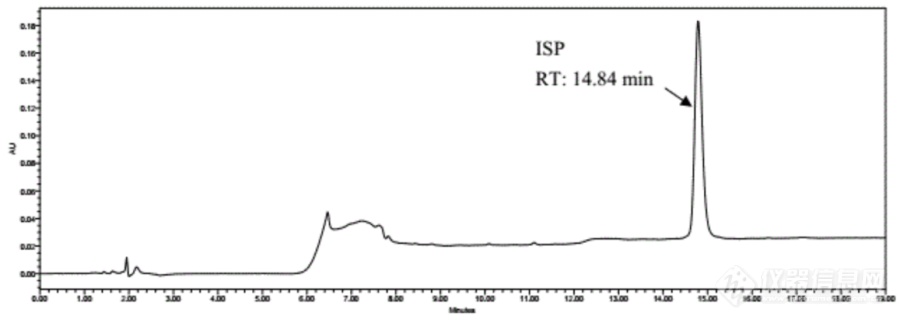
图1. ISP参考液相色谱曲线
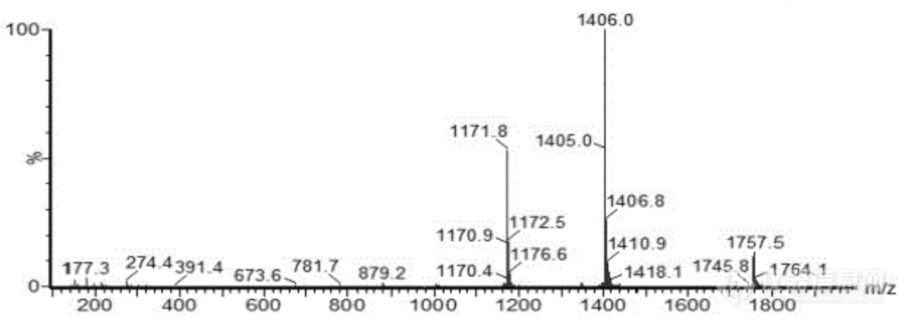
图2. ISP参考质谱图
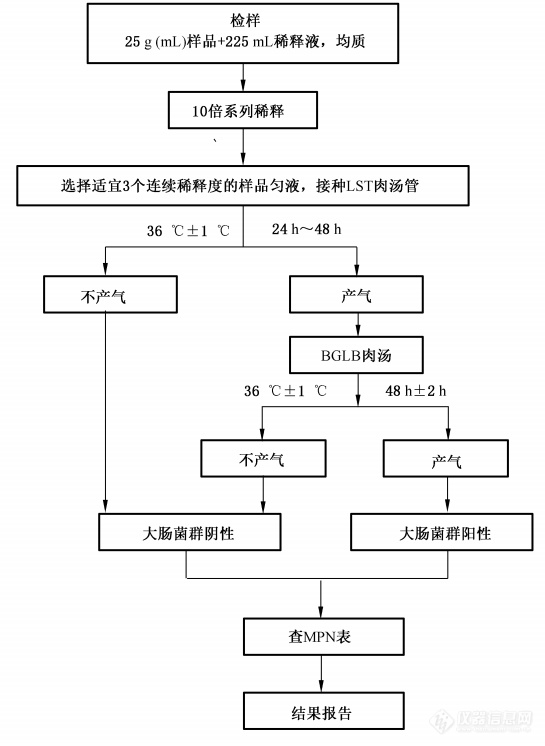
图3. 大肠菌群MPN计数法检验程序
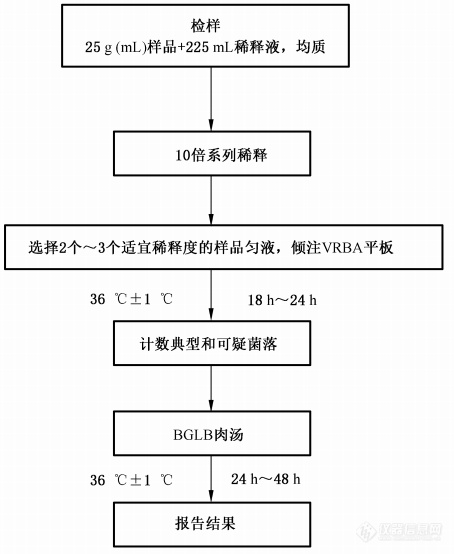
图4. 大肠菌群平板计数法检验程序
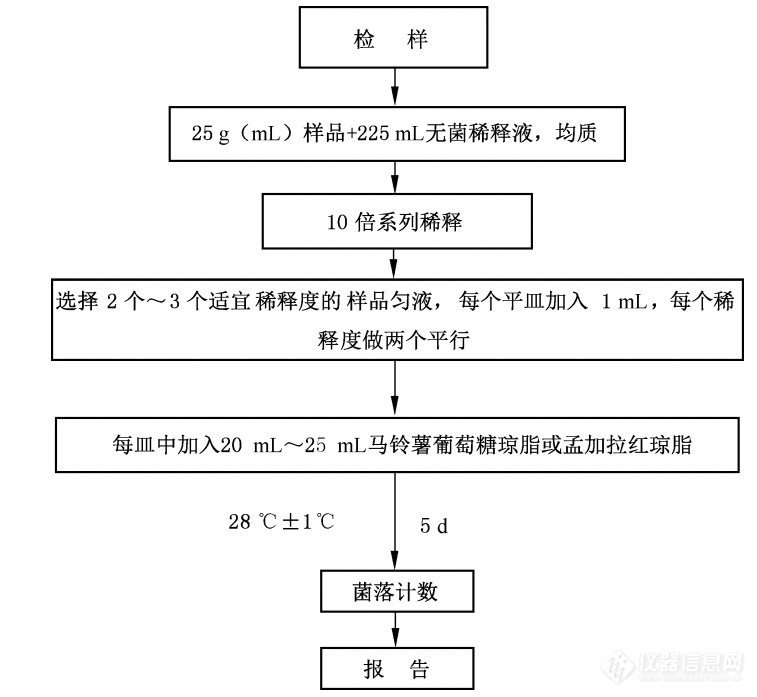
图5. 霉菌和酵母菌总数检验程序

图6. 沙门氏菌总数检验程序

图7. 单核细胞增生李斯特氏菌定性检验程序
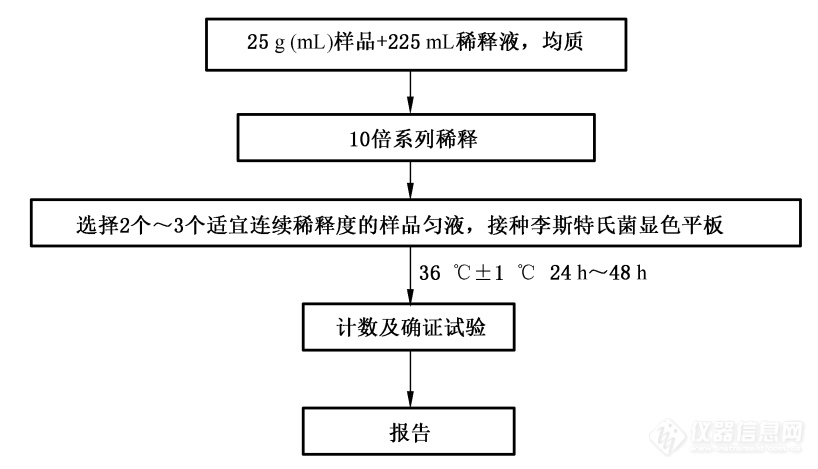
图8. 单核细胞增生李斯特氏菌平板计数程序
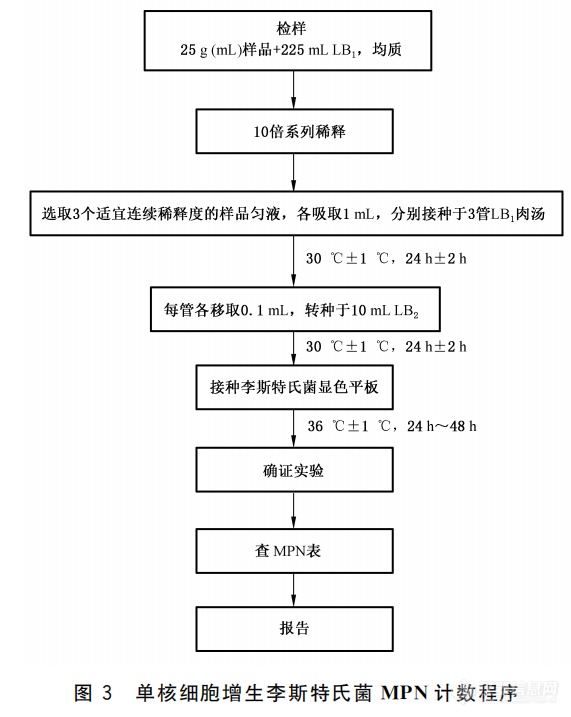
图9. 单核细胞增生李斯特氏菌MPN计数程序
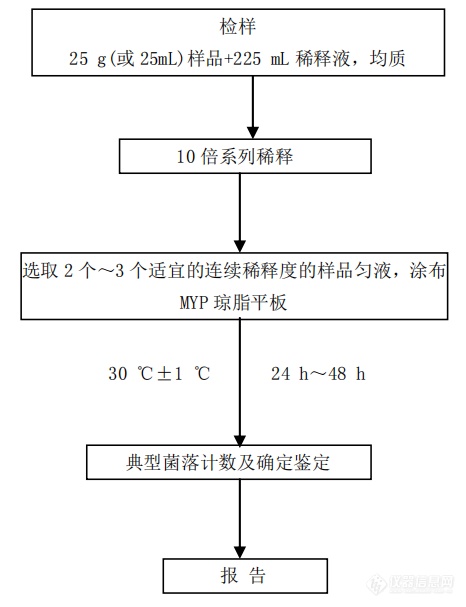
图10. 蜡样芽孢杆菌总数检验程序
参考文献
[1] | 樊凯凯,刘爱国,陈东等.冰结构蛋白及其应用研究[J].食品研究与开发,2012(09):221-225. |
[2] | RAYMOND JA, DEVRIESA L. Adsorption inhibition as amechanism of freezing resistance in polar fishes[J].Proc Natl Acad Sci,1977,74(6): 2589-2593 |
[3] | Jia Z,Davies P L.Antifreeze proteins:an unusual receptor-ligand interaction[J].Trends Biochem Sci,2002,27(2):101-106 |
[4] | Baderschneider B, Crevel R W R, Earl L K, et al.Sequence analysis and resistance to pepsin hydrolysis as part of an assessment of the potential allergenicity of Ice Structuring Protein Type III HPLC 12 [J]. Food and chemical toxicology,2002,40:965-978 |
[5] | C Bindslev -Jensen, E Sten, L K Earl,et al. Assessment of the potential allergenicity of ice structuring protein type III HPLC 12 using the FAO/WHO 2001 decision tree for novel foods[J]. Food and chemical toxicology,2003,41:81-87 |
[6] | 黄卫宁,邹奇波,潘振兴,等.冰结构蛋白对长期冻藏冷冻面团抗冻发酵特性与超微结构的影响[J].食品科学,2008,29(08):39-42 |
[7] | Warren C J,Mueller G M,McKown RL.Ice Crystal Growth Suppression Polypeptides and Methods of Preparation [P].US Patent 5,1992,118: 792-794 |

































