推荐厂家
暂无
暂无



 400-886-5615
400-886-5615
 留言咨询
留言咨询
 400-807-5250
留言咨询
400-807-5250
留言咨询
 400-860-5168转0600
留言咨询
400-860-5168转0600
留言咨询



基因组(genome)包含的遗传信息经转录产生mRNA,一个细胞在特定生理或病理状态下表达的所有种类的mRNA称为转录子组(transcriptome)。很显然,不同细胞在不同生理或病理状态下转录子组包含的mRNA的种类不尽相同。mRNA经翻译产生蛋白质,一个细胞在特定生理或病理状态下表达的所有种类的蛋白质称为蛋白质组(proteome)。同理,不同细胞在不同生理或病理状态下所表达的蛋白质的种类也不尽相同。蛋白质是基因功能的实施者,因此对蛋白质结构,定位和蛋白质-蛋白质相互作用的研究将为阐明生命现象的本质提供直接的基础。生命科学是实验科学,因此生命科学的发展极大地依赖于实验技术的发展。以DNA序列分析技术为核心的基因组研究技术推动了基因组研究的日新月异,而以基因芯片技术为代表的基因表达研究技术为科学家了解基因表达规律立下汗马功劳。在蛋白质组研究中,二维电泳和质谱技术的黄金组合又为科学家掌握蛋白质表达规律再铸辉煌。蛋白质组学(proteomics)就是指研究蛋白质组的技术及这些研究得到的结果。蛋白质组学的研究试图比较细胞在不同生理或病理条件下蛋白质表达的异同,对相关蛋白质进行分类和鉴定。更重要的是蛋白质组学的研究要分析蛋白质间相互作用和蛋白质的功能。蛋白质组学的研究内容包括:1.蛋白质鉴定:可以利用一维电泳和二维电泳并结合Western等技术,利用蛋白质芯片和抗体芯片及免疫共沉淀等技术对蛋白质进行鉴定研究。2.翻译后修饰:很多mRNA表达产生的蛋白质要经历翻译后修饰如磷酸化,糖基化,酶原激活等。翻译后修饰是蛋白质调节功能的重要方式,因此对蛋白质翻译后修饰的研究对阐明蛋白质的功能具有重要作用。3.蛋白质功能确定:如分析酶活性和确定酶底物,细胞因子的生物分析/配基-受体结合分析。可以利用基因敲除和反义技术分析基因表达产物-蛋白质的功能。另外对蛋白质表达出来后在细胞内的定位研究也在一定程度上有助于蛋白质功能的了解。Clontech的荧光蛋白表达系统就是研究蛋白质在细胞内定位的一个很好的工具。4.对人类而言,蛋白质组学的研究最终要服务于人类的健康,主要指促进分子医学的发展。如寻找药物的靶分子。很多药物本身就是蛋白质,而很多药物的靶分子也是蛋白质。药物也可以干预蛋白质-蛋白质相互作用。在基础医学和疾病机理研究中,了解人不同发育、生长期和不同生理、病理条件下及不同细胞类型的基因表达的特点具有特别重要的意义。这些研究可能找到直接与特定生理或病理状态相关的分子,进一步为设计作用于特定靶分子的药物奠定基础。不同发育、生长期和不同生理、病理条件下不同的细胞类型的基因表达是不一致的,因此对蛋白质表达的研究应该精确到细胞甚至亚细胞水平。可以利用免疫组织化学技术达到这个目的,但该技术的致命缺点是通量低。LCM技术可以精确地从组织切片中取出研究者感兴趣的细胞类型,因此LCM技术实际上是一种原位技术。取出的细胞用于蛋白质样品的制备,结合抗体芯片或二维电泳-质谱的技术路线,可以对蛋白质的表达进行原位的高通量的研究。很多研究采用匀浆组织制备蛋白质样品的技术路线,其研究结论值得怀疑,因为组织匀浆后不同细胞类型的蛋白质混杂在一起,最后得到的研究数据根本无法解释蛋白质在每类细胞中的表达情况。虽然培养细胞可以得到单一类型细胞,但体外培养的细胞很难模拟体内细胞的环境,因此这样研究得出的结论也很难用于解释在体实际情况。因此在研究中首先应该将不同细胞类型分离,分离出来的不同类型细胞可以用于基因表达研究,包括mRNA和蛋白质的表达。LCM技术获得的细胞可以用于蛋白质样品的制备。可以根据需要制备总蛋白,或膜蛋白,或核蛋白等,也可以富集糖蛋白,或通过去除白蛋白来减少蛋白质类型的复杂程度。相关试剂盒均有厂商提供。蛋白质样品中的不同类型的蛋白质可以通过二维电泳进行分离。二维电泳可以将不同种类的蛋白质按照等电点和分子量差异进行高分辨率的分离。成功的二维电泳可以将2000到3000种蛋白质进行分离。电泳后对胶进行高灵敏度的染色如银染和荧光染色。如果是比较两种样品之间蛋白质表达的异同,可以在同样条件下分别制备二者的蛋白质样品,然后在同样条件下进行二维电泳,染色后比较两块胶。也可以将二者的蛋白质样品分别用不同的荧光染料标记,然后两种蛋白质样品在一块胶上进行二维电泳的分离,最后通过荧光扫描技术分析结果。胶染色后可以利用凝胶图象分析系统成像,然后通过分析软件对蛋白质点进行定量分析,并且对感兴趣的蛋白质点进行定位。通过专门的蛋白质点切割系统,可以将蛋白质点所在的胶区域进行精确切割。接着对胶中蛋白质进行酶切消化,酶切后的消化物经脱盐/浓缩处理后就可以通过点样系统将蛋白质点样到特定的材料的表面(MALDI-TOF)。最后这些蛋白质就可以在质谱系统中进行分析,从而得到蛋白质的定性数据;这些数据可以用于构建数据库或和已有的数据库进行比较分析。实际上像人类的血浆,尿液,脑脊液,乳腺,心脏,膀胱癌和磷状细胞癌及多种病原微生物的蛋白质样品的二维电泳数据库已经建立起来,研究者可以登录www.expasy.ch/www/tools.html等网站进行查询,并和自己的同类研究进行对比分析。Genomic Solution可以为研究者提供除质谱外的所有蛋白质组学研究工具,包括二维电泳系统,成像系统及分析软件,胶切割系统,蛋白质消化浓缩工作站,点样工作站等;同时还可以提供相关试剂和消耗品。LCM-二维电泳-质谱的技术路线是典型的一条蛋白质组学研究的技术路线,除此以外,LCM-抗体芯片也是一条重要的蛋白质组学研究的技术路线。即通过LCM技术获得感兴趣的细胞类型,制备细胞蛋白质样品,蛋白质经荧光染料标记后和抗体芯片杂交,从而可以比较两种样品蛋白质表达的异同。Clontech最近开发了一张抗体芯片,可以对378种膜蛋白和胞浆蛋白进行分析。该芯片同时配合了抗体芯片的全部操作过程的重要试剂,包括蛋白质制备试剂,蛋白质的荧光染料标记试剂,标记体系的纯化试剂,杂交试剂等。对于蛋白质相互作用的研究,酵母双杂交和噬菌体展示技术无疑是很好的研究方法。Clontech开发的酵母双杂交系统和NEB公司开发的噬菌体展示技术可供研究者选用。关于蛋白质组的研究,也可以将蛋白质组的部分或全部种类的蛋白质制作成蛋白质芯片,这样的蛋白质芯片可以用于蛋白质相互作用研究,蛋白表达研究和小分子蛋白结合研究。Science,Vol.293,Issue 5537,2101-2105,September 14,2001发表了一篇关于酵母蛋白质组芯片的论文。该文主要研究内容为:将酵母的5800个ORF表达成蛋白质并进行纯化点样制作芯片,然后用该芯片筛选钙调素和磷脂分子的相互作用分子。最后有必要指出的是,传统的蛋白质研究注重研究单一蛋白质,而蛋白质组学注重研究参与特定生理或病理状态的所有的蛋白质种类及其与周围环境(分子)的关系。因此蛋白质组学的研究通常是高通量的。适应这个要求,蛋白质组学相关研究工具通常都是高度自动化的系统,通量高而速度快,配合相应分析软件和数据库,研究者可以在最短的时间内处理最多的数据。

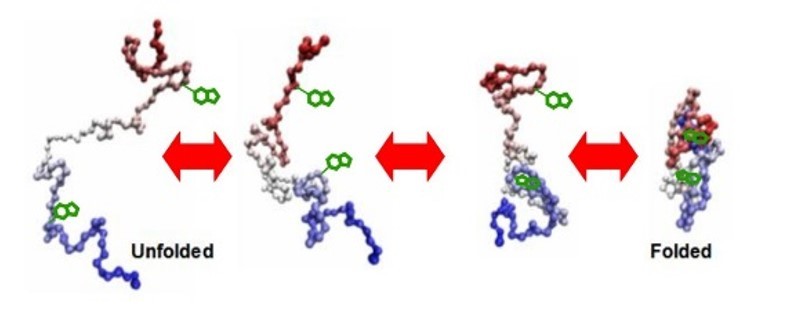
摘要:本文介绍了分子伴侣的基本概念,以及分子伴侣的几种主要类型;简要说明了蛋白质折叠的概念及特点;在此基础上,进一步阐述了分子伴侣的功能,并以GroEL和GroES为例简述了分子伴侣在蛋白质折叠过程中的作用机理。最后介绍了分子伴侣概念的延伸,及其研究意义和展望。关键词:分子伴侣 蛋白质折叠 折叠病 20世纪60年代,人们就发现了由于组成蛋白质的氨基酸错误可以导致分子病,后来人们发现,即使一级结构正常,蛋白质的二级结构乃至立体结构异常也可导致疾病,即蛋白质折叠病,如疯牛病、老年性痴呆、囊性纤维性炎等。蛋白质折叠病的发现激励人们去寻找蛋白质折叠的分子机理,近年来研究中发现,分子伴侣在在蛋白质折叠中起重要作用。1分子伴侣简介1.1分子伴侣的基本概念分子伴侣(Molecular Chaperone),也有人翻译为“分子伴娘”。1978年,Laskey等首先用“分子伴侣”描述核质素(nucleoplasmin)在核小体组装过程中的作用。1987年,Ellis将凡能促进蛋白质折叠和组装的蛋白质统称为分子伴侣。随后,Ellis等又提出了分子伴侣的基本概念:在蛋白质折叠和组装过程中,分子伴侣防止多肽链内或链间因疏水键等相互作用表面瞬间暴露而形成错误结构,并且还可以破坏已经形成的错误结构。分子伴侣本身不是折叠或组装产物的一部分。1.2分子伴侣的几个例子Nucleoplasmins:体内的一系列过程,如DNA复制,RNA转录与剪接,核小体或核糖体的装配,都涉及到带正电的蛋白质与带负电的核酸之间较强的离子键的相互作用。实验发现,这些过程都与Nucleoplasmin相类似的蛋白质的参与。Charperonin(Cpn):是指在细菌、线粒体、质体中发现的一类序列同源的Charperonins,该家族具有独特的双层7-9元环状结构的寡聚蛋白(Hemminngwen;cheng 1998),它们的作用是促进体内正常条件以及应急反应下的蛋白质折叠,这一过程需要ATP提供能量。Cpns包括细菌的GroEL、叶绿体的Rubisco亚基结合蛋白(RuSBP)与线粒体的热休克蛋白Hsp60。Stress-70家族:该家族首先在热休克反应中发现,并研究多年,近些年来,发现Stress-70也在蛋白质的折叠与装配过程中起作用,因而受到广泛关注。参与这些作用的Stress-70的成员有:E. coli的DnaK、酵母细胞质的Ssa1p和Ssa2p、内质网的Kar2p和线粒体的Ssc1p。哺乳动物细胞质的Hsp70蛋白和Prp73多肽识别蛋白、内质网的Bip。这些蛋白可被细胞内未折叠蛋白质的增多而诱导并识别靶分子,在其他热休克蛋白或细胞因子的参与下,水解ATP调节蛋白的构象或折叠状态。Stress-90家族:分子量在90ku左右,包括大肠杆菌胞
知道凯氏定氮法测定蛋白质含量的计算公式中,有个V3是吸取消化液的体积,这个体积是在什么时候添加的,怎么添加的。如果看到请帮忙解答一下,谢谢了









 我要推广仪器
我要推广仪器
 下载APP
下载APP